

Dominique.
-
Compteur de contenus
1 277 -
Inscription
-
Dernière visite
Messages posté(e)s par Dominique.
-
-
Film biologique Chilomonas
Dans le jardin il y a un seau qui me sert à transporter les mauvaises herbes – la chaux - les fruits avariés et toutes sortes de choses Ce seau est resté sans emploi durant un certain temps - se remplissant doucement de l’ eau de pluie - ( mais pas depuis 15 jours car dans ce coin de Normandie la sécheresse s’ est installée )
Il y a quelques jours est apparu un film orange à sa surface.

Ce milieu turpide révèle un Ph à 8,5 soit basique
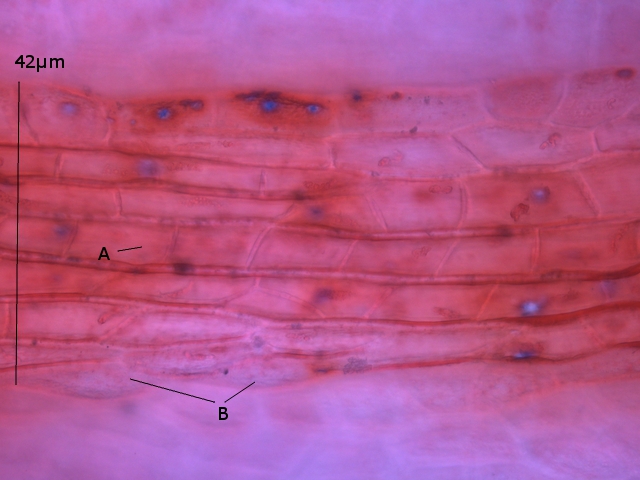
La face d’ ‘ une lame est posée sur la surface. Celle-ci est examinée au X 100
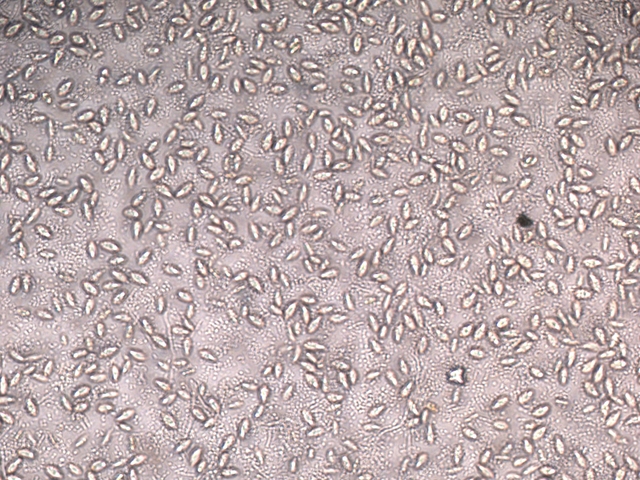
Sur cette photo on constate la présence de deux types d éléments dominants
--A -la présence de protistes qui sont extrémement mobiles
--B-- la présence de microbes
Le protiste
Examen au X 600 - ( le problème est que pour les forts grossissements tout ce petit monde bouge beaucoup ).
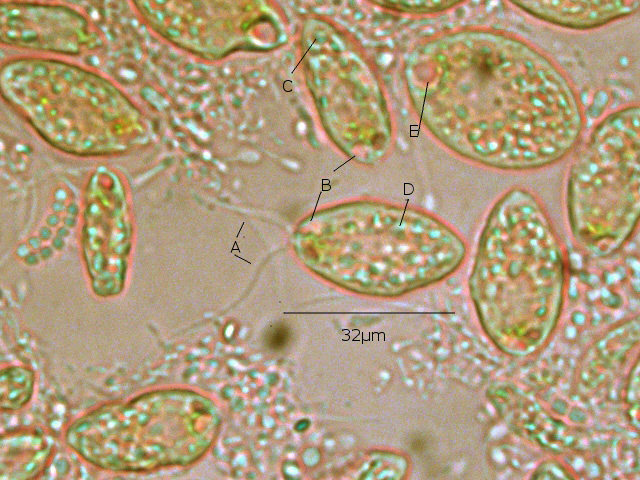
Taille 32µm de long
A - Ce protiste est porteur de deux flagelles à situation apicale
B - A la base de ces flagelles il existe une dépression le cytopharynx
C - Le noyau est situé dans la partie postérieure
D Le cytoplasme est bourré de grains ( d’ amidon ?)
E Il existe une vacuole contractile
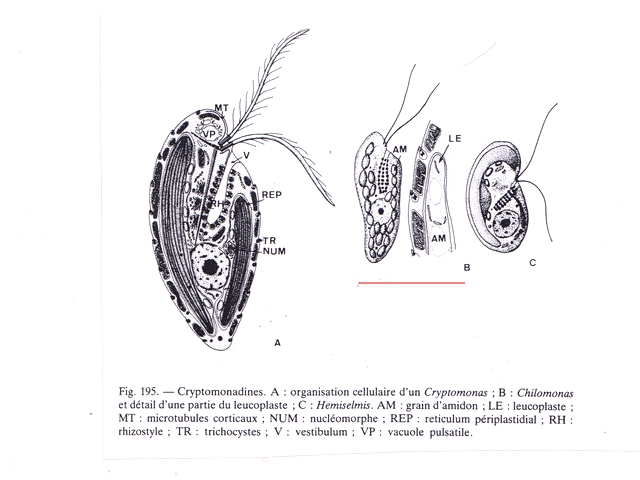
Cette description peut correspond au genre Chilomonas
Dans le précis de Protistologie on peut lire « les plastes sont pariétaux de couleur souvent vert olive mais parfois brune , rouge ou même bleue » ce qui explique la coloration brune orangée du film ( l’eau en dessous du film est transparente ) .
Classification:
Empire: Eukaryota
Kingdom: Chromista
Subkingdom:Hacrobia
Phylum Cryptista
Subphylum: Rollomonadia
Class: Cryptophyceae
Order :Cryptomonadales
Family: Cryptomonadaceae
Genus :Chilomonas
Les microbes
Il est réalisé une coloration de Gram : technique violet de Gentiane 30 secondes - rincer - Lugol 20 secondes - rincer - Alcool 95 pour décolorer 5 à 10 secondes –Fuscine 30 secondes -
Séchage à 40 °C Examen X 900
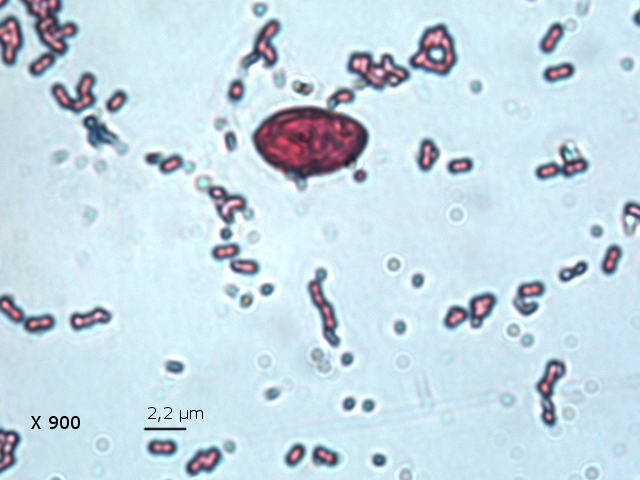
On obtient un résultat Gram négatif pour un microbe de type coccus associés par 2( Diplocoque ) .
Conclusion :
Les films biologiques sont en fait des milieux complexes - dans celui-ci il y a deux espèces dominantes Un diplocoque gram négatif et un protiste du genre Chilomonas . Il y a probablement d’ autres espèces mais elles ne sont pas facilement identifiables en raison du surpeuplement par la population de cocci et de protistes
. Une autre observation avait déjà été faite de ce phénomène de film biologique avec pour peuplement dominant une algue https://forum.MikrOscOpia.com/topic/13864-algue-sur-une-mare/?hl=%2Bfilm+%2Bbiologique
Références:
https://eol.org/pages/49278345
Précis de protistologie de Pierre de Puytorac et coll edition Boubée 1997 – ( on prend conscience de la puissance des études qui ont été faite au 20éme siècle sur les protistes ( surtout en microscopie électronique ) – mais pour un amateur ce livre est illisible
Dominique.
dd
-
Bonsoir Tryphon
Je ne contredis pas - ton hypothèse en regard de la publication me semble seulement avoir une probabilité modeste . Mais cette situation est difficile devant l' accumulation de beaucoup d' imprécisions . .
Amicalement
Dominique
-
Bonsoir Jean -Marie et Tryphon
Tryphon ton hypothèse est , en effet, correcte mais n' est pas acceptable si on se réfère au site suivant
https://player.slideplayer.com/15/4736248/#
il faut cliquer sur l'image pour les faire défiler
Merci Jean - Marie pour l 'information . D'aprés cette publication on pourrait le classer parmi l' espéce Diphtherophora lata
Amicalement
Dominique
-
Bonsoir Jean - Marie
Connais tu un site pour la reconnaissance des vers ? Je sais que l' image du pharynx est un élément important pour la détermination ( en agrandissant l' image on pourrait y trouver les indices nécessaires ) .
Amicalement
Dominique.
-
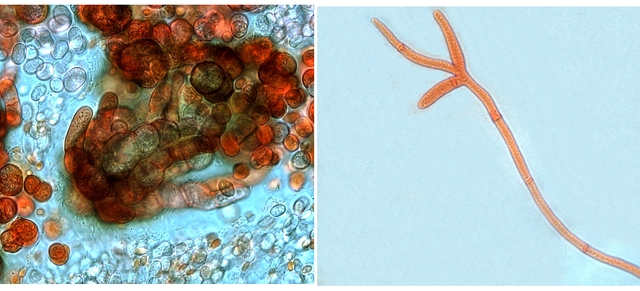
Les hyphes sont septés ( c’est à dire cloisonnés entre deux cellules fongiques ).
On constate des cellules conidogénes ( production de spores directemement d 'une cellule d' hyphe .
Ces aspects sont compatibles avec une moisissure du genre Microdochium .
Cette moisissure est bien connue des cultivateurs puisqu’ elle fait partie des agents responsables de la Furasiose .Il s’agit d’ une maladie cryptogamique du blé . La fusariose des épis est provoquée par plusieurs espéces de fungus appartenant aux genres Microdochium et Fusarium
L’infection entraîne souvent le blanchiment de tout ou partie de l’épi . La fusariose qui affecte l’épi peut entraîner des pertes de rendement pouvant aller de 30 à 70 %. Ces pertes sont dues à la nécrose précoce des épis et à des grains mal remplis et déformés;
Une diminution de la qualité boulangère et du taux de germination. Les fusarioses sont à l’origine des mycotoxines.
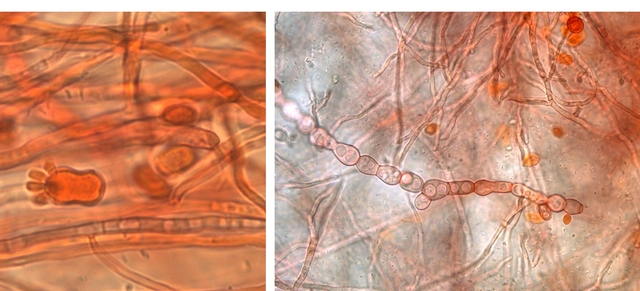
Prélèvement dans la partie profonde
Ce prélèvement met en évidence la présence d’une grande quantité de vers
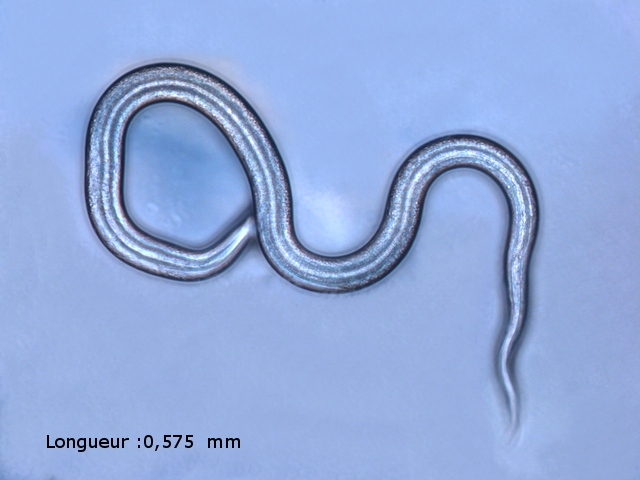
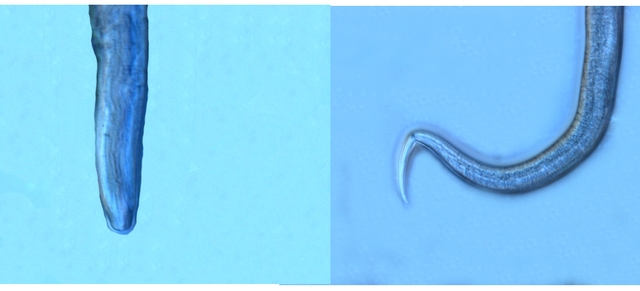
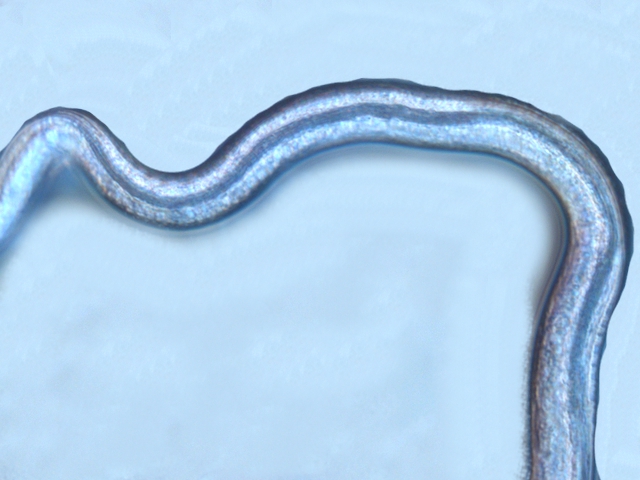
Il existe un vers plat de plus grande taille qui se déplace sur la lame - il y en a très peu .
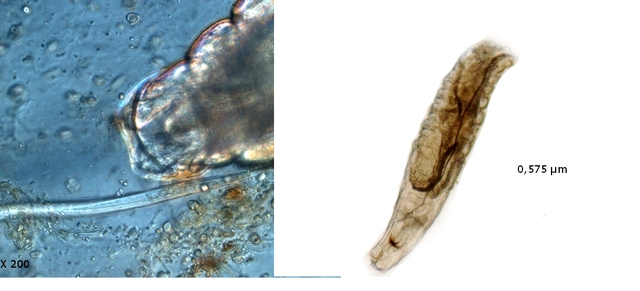
sur la petite photo de droite on met en évidence la différence de taille entre les deux espèce -
il y a une erreur sur le photo de droite : le vers plat fait 0,950 mm (soit une longueur un peu plus longue que le vers rond )
Je ‘n’ ai pas trouvé le nom de ces deux espèces .
Les autres moisissures qui se sont développées sur le milieu de culture .
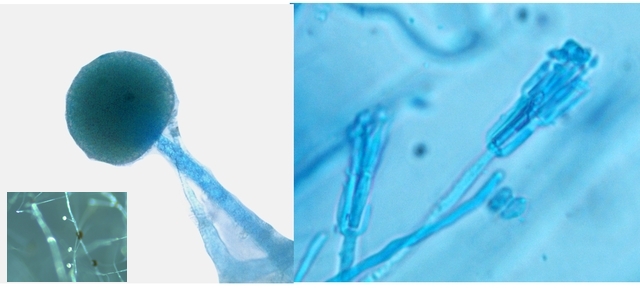
A gauche - Sporange et Sporangiophore de Rhizopus ( en bas à gauche examen en episcopie )
A droite - Pénicillium
Après 15 jours d’ évolution l’aspect de la pêche a changé - la couche de moisissures a disparu ou s’ est considérablement atténuée - par contre il existe une couverture grise feutrée dont on fait le prélèvement .
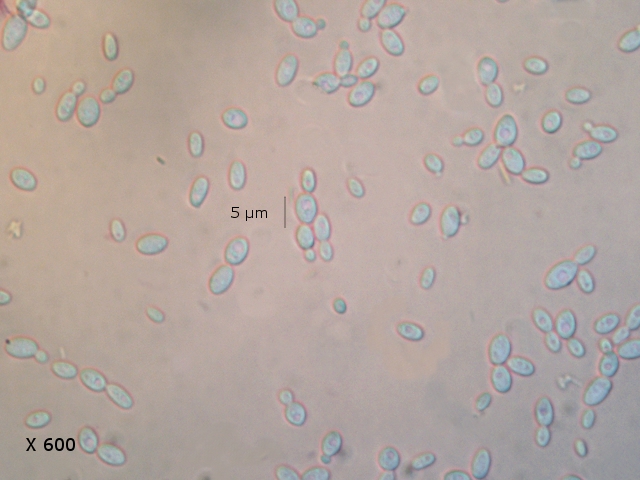
Il s’agit d’ une colonie de Levures dont le développement a été permis par les modifications des tissus de la pomme sous l’effet de la moisissure et des bactéries .
Enfin on constate comme à chaque fois qu’un fruit est laissé à pourrir au grand air le développement de drosophiles avec la présence à la base de la pêche d’un grand nombre de pupes
https://forum.MikrOscOpia.com/topic/18780-trompe-mouche-drosophile/?hl=drosophile

La pourriture d’un fruit devient un microcosme pour les saprophytes qui trouvent leur carbone dans la matière organique morte Ce monde est surtout dominé par les moisissures et les bactéries - J’ ai laissé le milieu évoluer - Après 15 jours le Penicillium et le Rizopus se sont installés en maîtres . puis ont quasi disparu pour être remplacés par une vaste colonie de levures - Après 15 jours les drosophiles sont toujours là.
Ref
Czapek https://fr.sperohope.com/agar-czapek-fundamento
Fusariose du blé
https://mycolab.pp.nchu.edu.tw/rice_fungi/species_info.php?species_name=Microdochium%20poae
https://www.gene.affrc.go.jp/databases-micro_images_detail_en.php?id=30774
Dominique .
-
Pourrissement : le cas d’une pêche - les agents responsables
Dans un coin du garage une pêche n’ a pas été emportée à la cuisine - Elle a subi son processus de dégradation habituel
Mais quels sont les agents responsables de cette dégradation en dehors de l’inévitable prolifération bactérienne .
Prélèvement de la couche superficielle :

Le premier aperçu est celui d’une prolifération de moisissures - Les colonies bleutées sont habituelles sur les fruits en cours de pourrissement .Ce sont des colonies de Penicillium -mais quel est le nom de la colonie marron et noire ? .
Un prélèvement local ne donne souvent pas de solution exploitable ( images microscopiques trop confuses ) d’où la nécessité de faire une culture .Un milieu de Czapek est fabriqué
Les boîtes de Petri ensemencées avec un milieu dilué au 100 ( prélèvement d’un fragment mis dans un flacon de 10cc -prélèvement de 1 c dans 10 c - 2 fois de suite ) Étalement par gouttes sur le milieu et mise à l’ étuve à 25 °c :résultat 48 heures après
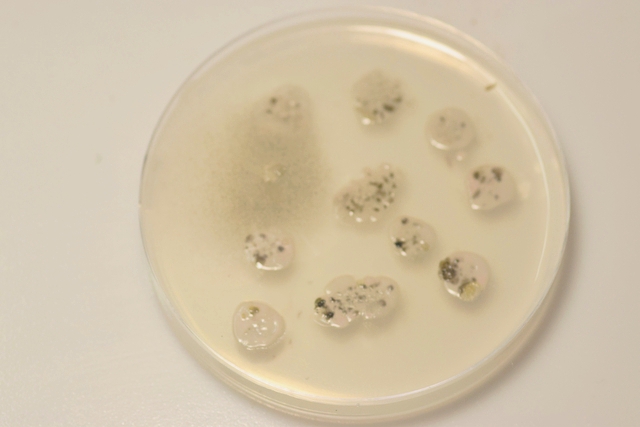
Dans chaque goutte on constate le développement de colonies noires -A noter en haut à gauche une colonie de Rhizopus de croissance extrêmement rapide .et envahissante .
Examen à la binoculaire de ces colonies noires :
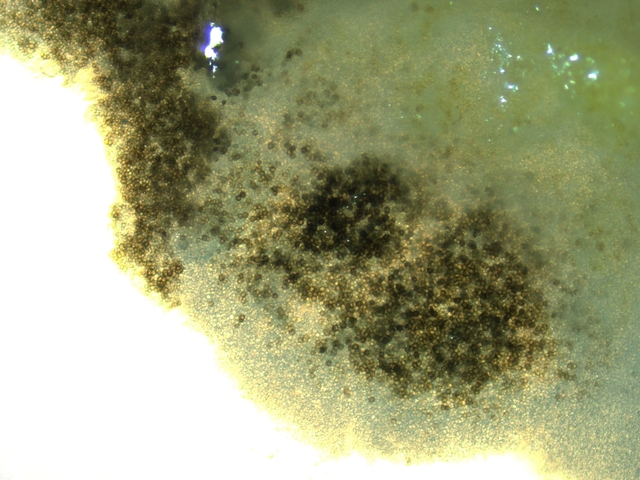
Prélèvement d’une colonie et étalement sur un lame - on fait une coloration au Rouge Congo Ammoniaqué.
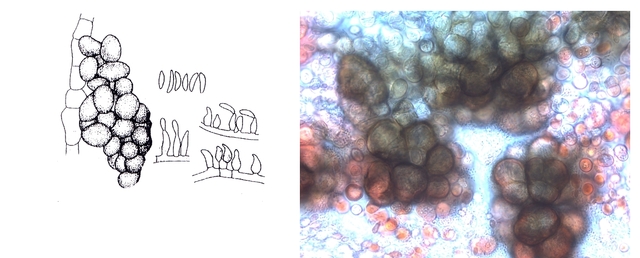
Le schéma initial résume les différents aspects que l’on va rencontrer en examinant la lame
Sur la photo de droite il est mis en évidence ce qui ressemble à des sporodochies ( ce sont des regroupements d'un agrégat de filaments et de conidiophores le tout ayant une forme de coussin.
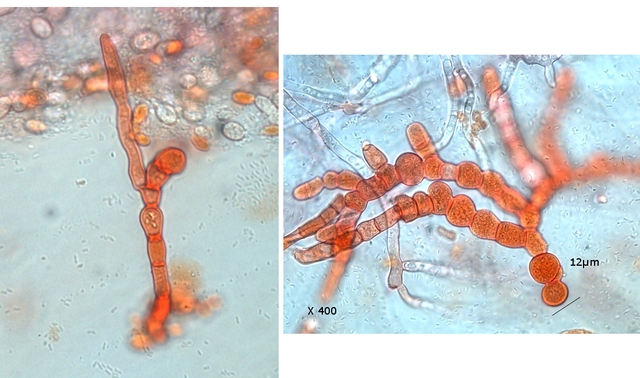
Les cellules sont souvent groupées en stromas qui forment des masses irrégulières de filaments végétatifs.
-
Polypore
Genre Trichaptum
Du fait de la sécheresse le sol de la forêt n’ est pas très riche en champignons - Les polypores par contre ne sont pas difficiles à trouver .Dans la rue du jardin botanique le tronc d’un vieux charme mort est toujours en place - Dessus existent plusieurs colonies de polypores . Ces colonies se développent depuis plusieurs années.
Habituellement l'apparition des sporophores des champignons est intermittente du fait des variations climatiques et leur durée de vie est souvent très courte (quelques jours) ; mais le vie de certains sporophores peur dépasser quinze ans chez les polypores coriaces comme c’est le cas pour le genre Trichaptum
Cette longue survie implique une structure qui puisse résister aux intempéries.

Exploration de la structure de ce Basodiomycéte .
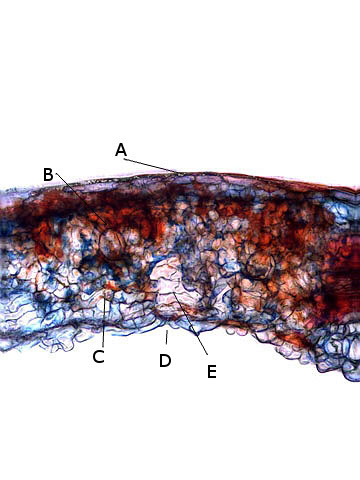
Coupe d’une langue de ce polypore

La coupe met donc en évidence 3 zones
Une zone d’ attache au tronc - d’où partent les hyphes qui vont coloniser le bois en cours de décomposition -
Une zone épaisse qui assure la rigidité et la fixation
Une zone fertile périphérique , située en position inferieure, où vont se développer le sporocystes
Une zone d’ attache au tronc - d’où partent les hyphes qui vont coloniser le bois en cours de décomposition -

Une zone épaisse qui assure la rigidité et la fixation ( X100 ).
Partie superieure :
Cette zone de coupe est située près de sa fixation sur l’ arbre – il est constaté que les hyphes semblent morts et qu’ils forment un amas compact du moins dans sa partie supérieure.
Coupe en dessous de la zone ci-dessus qui est la zone de circulation entre les hyphes qui pénètrent le bois et le sporophore

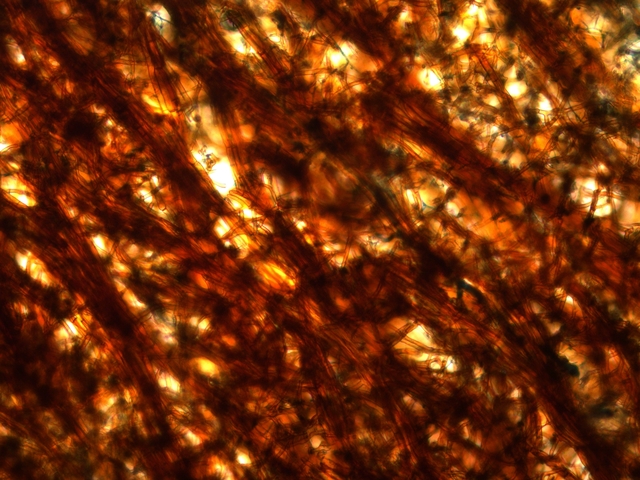
La structure est formée d’un emmêlement d’un grand nombre d’ hyphes.
La zone génératrice :Les lamelles sont situées sur la partie inferieure du sporophore :
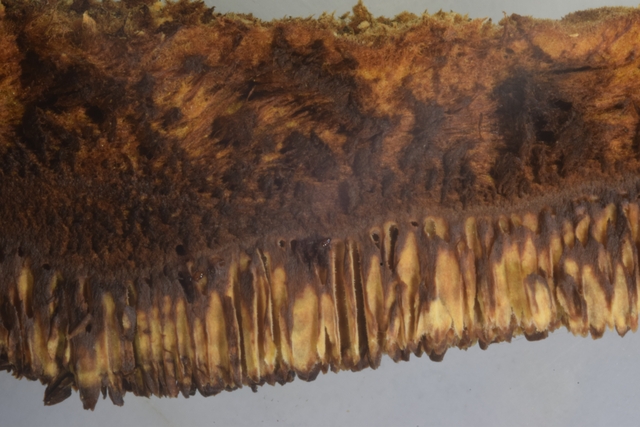
Vue de la face inferieure sur un sujet plus jeune que celui utilisé pour les coupes précédentes.

Coupe d’une lamelle X 100

Les zone fertiles sont donc situées de part et d’autre de la lamelle qui est formée par un compactage d’ hyphes :
Images précisant le développement reproducteur et la zone inter lamellaire dans sa partie distale .
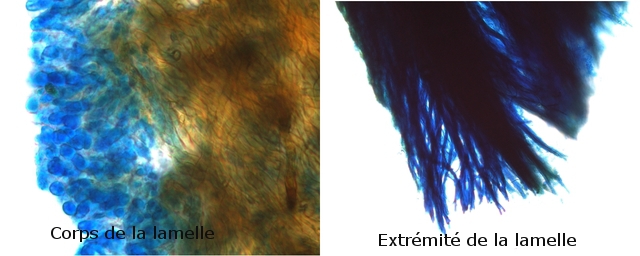
Les spores :
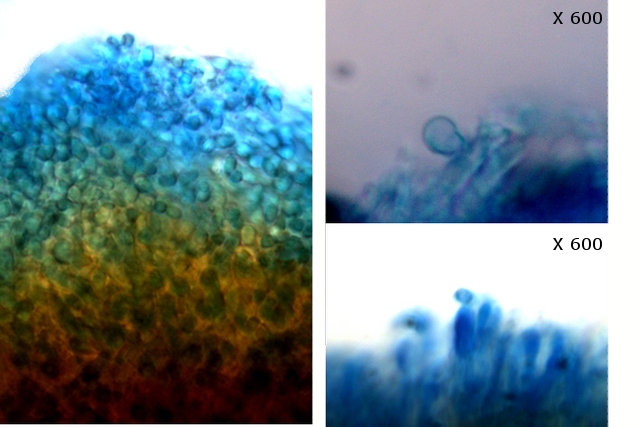
L’examen des coupes met aussi en évidence trois types de spores :
Les conidies ( ou spore asexuées
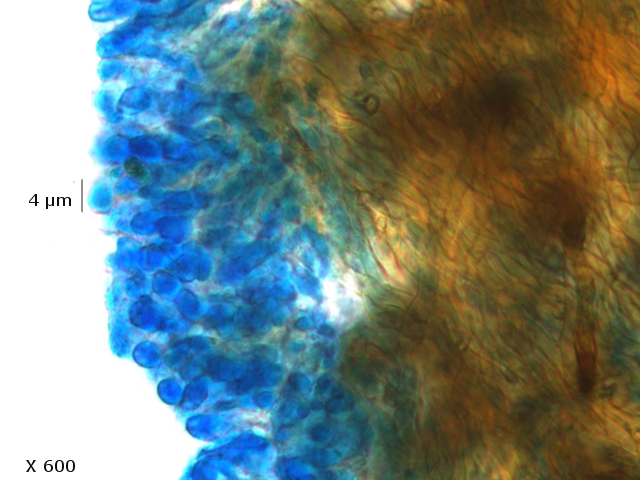
Le plan profond est formé par de très nombreux hyphes - Le plan superficiel est lui formé de structures cellulaires rondes /ovalaires plus ou moins régulièrement empilées . devant correspondre à des empilements de conidies (Une conidie (ou conidiospore) est une spore assurant la multiplication asexuée des champignons-).
Les chlamydospores ( spore asexuée de résistance ).
Les chlamydospores sont des spores asexuées de forme sphérique ou ellipsoïde, produites par certains champignons basidomycètes. Elles sont généralement plus grandes que les spores sexuelles (basidiospores) et sont entourées d'une paroi épaisse et résistante.
Les chlamydospores sont produites dans diverses conditions, notamment en réponse à des conditions défavorables, telles que la sécheresse, le froid ou la malnutrition. Elles permettent au champignon de survivre à ces conditions et de se propager lorsque les conditions redeviennent favorables.
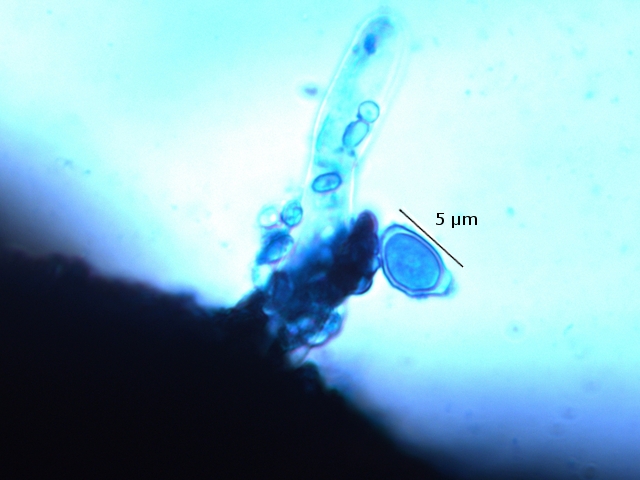
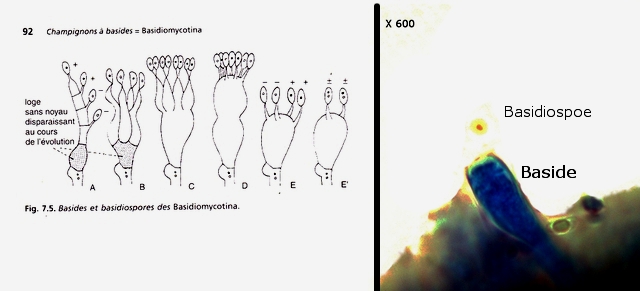
Les basidiospores .
Dans cette observation malgré un nombre important de coupes il a été très difficile de mettre en évidence des basides et encore moins des basides correctes surmontées par son image bien connue avec ses 4 spores .Cette forme sexuée de reproduction n’ est donc pas la forme dominante .
Ces spores sont en effet la forme sexuée de la propagation du champignon .
Ces spores sont en effet la forme sexuée de la propagation du champignon .
La baside sur la droite de la photo est de qualité plus que modeste , elle est porteuse d’une seule basidiospore et le stèrigmate qui la lie à la baside n’est pas visible .
Les basidiospores sont formés par l’union des deux noyaux haploïdes ( n chromosomes ) des cellules d'hyphes de polarités différentes ( on ne parle pas de sexe ) qui convergent deux à deux, ces deux noyaux coexistent d’abord sans caryogamie puis s’unissant forment un Zygote à 2n chromosomes

Ce zygote va subir une méiose qui va donner naissance à 2 noyaux haploïdes ( n chromosomes ) En fonction du nombre de mitoses qui vont suivre on va obtenir 2 ou 8 spores - ( mais beaucoup ne se développeront pas ).
Kingdom: Fungi
Division: Basidiomycota
Class: Agaricomycetes
Order: Hymenochaetales
Family: Incertae sedis
Genus: Trichaptum
Murrill (1904)
Référence :
Les champignons mycologie fondamentale et appliquée de P Boucher et col editeur Masson
Dominique
-
Bonsoir Jean -Luc , Jean-Marie , tous.
Les coupes ont été faites après avoir placé l' élytre sans préparation dans un moule où j' ai coulé de la paraffine - le bloc a été coupé au Microtome de Minot 70 µm d' épaisseur-
Le mécanisme d' ouverture des élytres est musculaire seul l' aile s 'ouvre par pression hydraulique ( hemolymphatique ).
Amicalement
Dominique.
-
Coléoptère élytres ailes
Lors d’une marche dans la forêt de Balleroy ( 14 ) , on découvre , gisant sur le sol une lucane cerf-volant femelle - avec une élytre entre ouverte .

Les coléoptères ont deux paires d’ ailes - 2 ailes du plan supérieur ou élytres - et 2 ailes du plan inférieur membraneuses .
Au toucher l’élytre est rigide - franchement dur - non pliable .Une coupe de cette élite est réalisée :
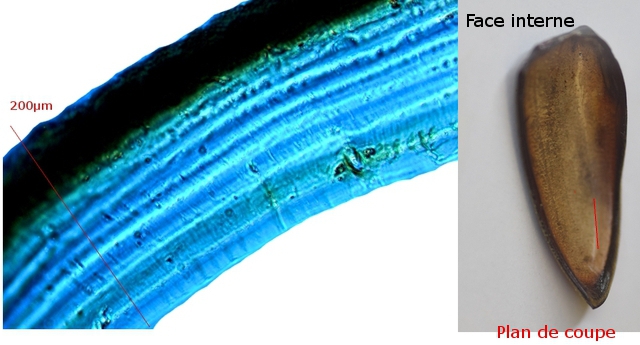
La résistance de cet organe tient à sa structure - Il est constitué de plaques comme sont réalisées les poutres en lamellé-collé dont la résistance est telle qu’ elles permettent des portées de 45 mètres ;
Image en lumière polarisée .
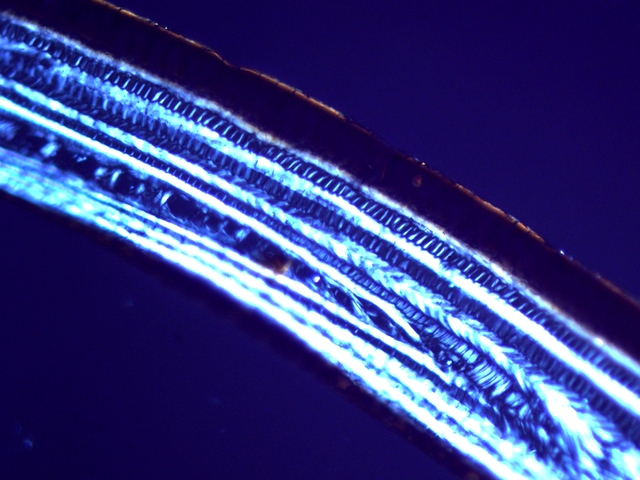
L’aspect est celui d’un empilement cellulaire qui s’organisent par plaques .
Les élytres sont constituées - comme le reste du squelette externe des insectes de chitine - Cette protéine est renforcée par la présence de calcium et de phosphore
L'élytre des coléoptères est constituée de trois couches cellulaires principales :
-- L'épicuticule est la couche externe de l'élytre. Elle est composée de chitine et de lipides. Elle est imperméable à l'eau et à l'air.
--L'endocuticule est la couche intermédiaire de l'élytre. Elle est composée de chitine et de protéines. Elle est plus flexible que l'épicuticule.
--L'hypoderme est la couche interne de l'élytre. Elle est composée de cellules vivantes. Ces cellules produisent la chitine et les autres substances qui constituent l'élytre.
En plus de ces trois couches cellulaires principales, l'élytre contient également des poils, des glandes et des cellules sensorielles
Les glandes sont présentes dans l'hypoderme. Elles produisent des substances telles que de la cire, des phéromones et des toxines.
Les élytres protègent deux ailes membraneuses : Ces ailes membraneuses ont 2 positions :
Repliées :

Ouvertes :

Comment ces deux ailes pliées peuvent elles se déployer ? ( elles le font avec une très grande rapidité comme on peut facilement le constater quand on assiste à l‘ envol d’une coccinelle ).
L’aile est pourvue de deux articulations :
Articulation sur le bord d’attaque ( X 40 + recadrage).

Articulation au milieu de l’ aile sur une grosse nervure ( X 40 +recadrage )

Par quelle mécanisme ce déploiement des ailes peut-il se faire Puisque il n’est pas possible de mettre en évidence la présence ni de muscle , ni de tendon ?
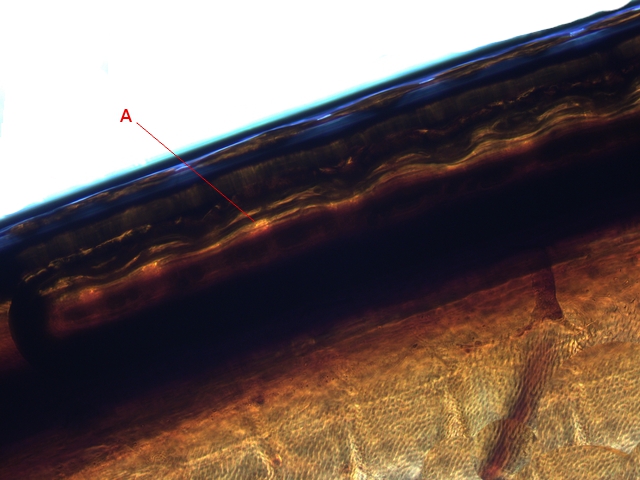
En réalité l’examen bord des nervures semble apporter la solution :
Articulation antérieure : photo prise le long du bord d’attaque :
Articulation du milieu de l’ aile le long d’une nervure :
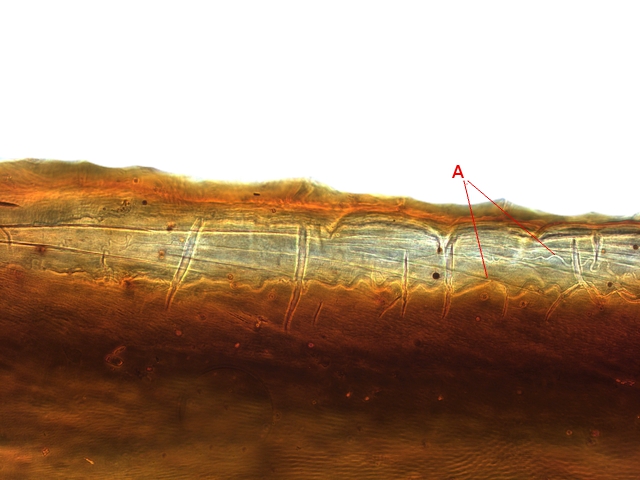
Sur ces deux photos au X 100 il est mis en évidence la présence d’un système vasculaire - Le moteur du mouvement des ailes est donc de type hydraulique à l’ image des bras des gros instruments de travaux publiques - L’ hémolymphe comprimé par les muscles thoraciques va conduire à une modification de la forme des articulations dont la conséquence sera l’ ouverture de l’ aile ( ce qui explique que les articulations ne sont pas comme celle de notre fémur avec le bassin ou de notre humérus avec l’ omoplate qui pour fonctionner nécessité la traction de tendons ) Ici c’est la déformation des surfaces articulaires qui va créer le mouvement .
Le mouvement des élytres par contre se déplie pas activité des muscles et des tendons thoraciques .
Régne Animalia
Embranchement Arthropoda
Sous-embr. Hexapoda
Classe Insecta
Sous classe Pterygota
Infra classe Neoptera
Super ordre Endopterygota
Ordre Coleoptera
Sous-ordre Polyphaga
Infra-ordre Scarabeiformia
Famille Lucanidae
Genre Lucanus
Espéce Lucanus cervus
Quasi menacé
Dominique
-
Bonjour Jean-Marie
Je pense que l’ on peut placer ces plantes dans un jardin public à condition de les enlever dès que les fruits sont mûres .Cela nécessite donc une surveillance Je vais faire attention à l’ évolution de ces plants -( sinon je signalerai à la municipalité le risque et la nécessité de les enlever dès la fin aout - période ou les graines ne sont pas encore expulsées ) - - Ces plants ont-ils une toxicité moindre ? - il faudra se renseigner .
Amicalement
Dominique
-
Ricin commun
Ricinus communis
Cette année la municipalité a décidé de placer des plants de Ricin sur les massifs du parc public-le résultat est très agréable :cette plante par son feuillage et l’ aspect inhabituel de ses feuilles donne une impression d’ exotisme dans ce petit port normand
.

Les fleurs sont regroupées ( Formant des cyathes - caracteristiques des euphorbiacés dont fait partie le Ricin commun
Les fleurs mâles et les fleurs femelles sont portées par la même plante ( La plante est dite monoïque).
Elle fleurit avec le début de l’ été .
Je propose de regarder cette plante au travers du microscope
La feuille
La tige
La fleur mâle
La fleur femelle - la graine
La feuille
Les feuilles sont palmées (avec 5 à 7 lobes pointus), dentelées, vertes avec des nervures plus ou moins rouges. Elles sont reliées à la tige principale par de longs pétioles (30 centimètres).

Coupe de la feuille à hauteur d’une nervure
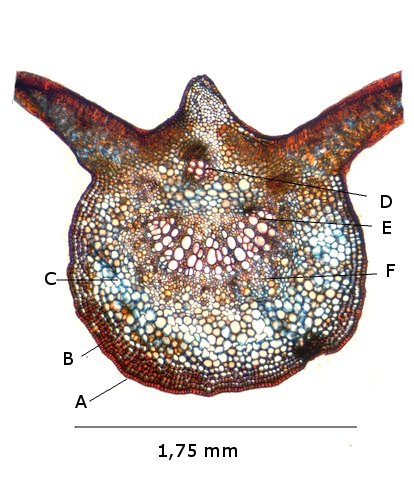
A – Epiderme
B – Periderme (Phelloderme -phellogène)
C – Zone qui contient de la périphérie vers le centre : les cellules de collenchyme – les cellules parenchymateuses - le cellules de sclérenchyme
D – Xyléme primaire
D – Xyléme secondaire
E – Cambium
Coupe d’une feuille à hauteur du limbe
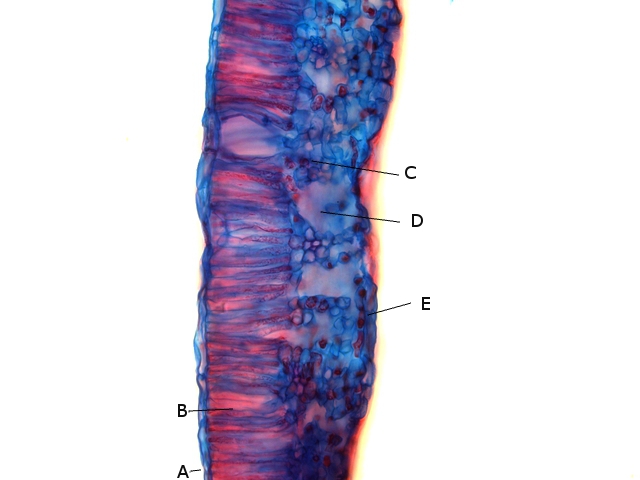
A –Cuticule
B --Cellules palissadiques
C –Cellules du mésophile
D –Lacunes aériennes ( contenant l’ oxygène produit par le processus de photosynthèse et le CO2 d’origine extérieur )
E –Epiderme inferieur
La tige principale
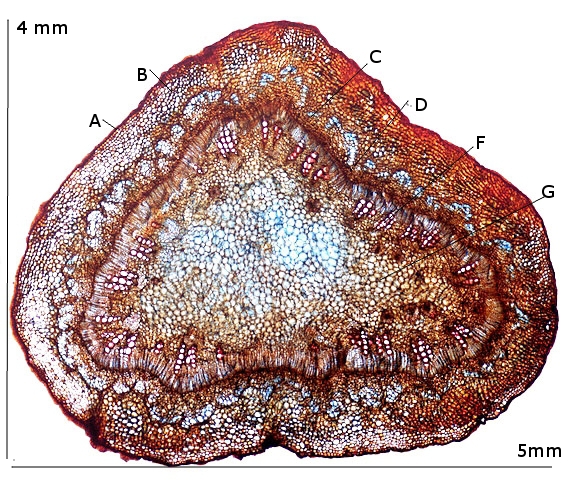
A – Epiderme
B – Periderme (assise subéro-phellodermique ou cambium subéro-phellodermique, est un méristème secondaire cortical )
C –,Endoderme correspond à l’extrémité du cylindre central d’un faisceau libéro-ligneux– Phloeme secondaire
D – Cambium périphérique
F – Xyléme
G –Cellules médullaires
La tige porte des fleurs mâles et femelles

A— Fleur mâle ouverte avec son bouquet d’étamines.
B -- La fleur femelle a ici laissé la place aux fruits en cours de développement.
Les fleurs mâles sont groupées le long de la tige :

L’ efflorescence des petites fleurs mâles , (Cyathe de fleurs mâles )- Par contre dès leur ouverture elle développent des bouquets blancs
Ces étamines ont la particularité de former des arborescences par division dichotomique des filets ; L’anthère est fixée à chaque fin de division dufilet de l'étamine par le « connectif »,
Elle est composée de deux thèques possédant chacun deux sacs polliniques avant maturité, nombre constant chez les Angiospermes. Ceux-ci fusionnent à maturité pour donner deux loges (anthère biloculaire),. Elles s'ouvrent, généralement par déhiscence, pour répandre les grains de pollen à l'extérieur.
L’organisation des étamines:

Les anthères:

La dispersion se fait par le vent .
Le pollen:
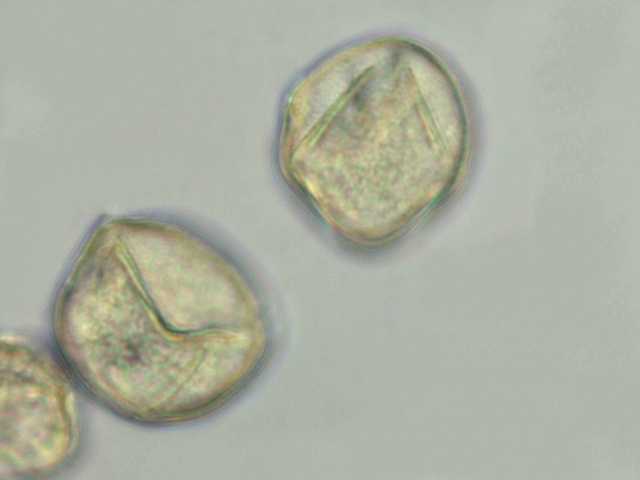
Taille des pollens autour de 50 µm .
Les fleurs femelles
Elles possèdent 5 tépales
Elles forment des inflorescences très rouges.
La fleur du ricin commun a 3 carpelles. Fusionnés qui forment le Gynécée.
L’ensemble des carpelles forme le pistil central surmonté de 3 stigmates qui sont bifides
L'ovaire contient 3 ovules,
Photo des 3 stigmates :

Les fruits
Ce sont des capsules tricoques :
Ces fruits sont l’un des éléments les plus spectaculaires de la plante .Leur péricarpe rouge est très ornementé de longues pointes .

La coupe d’une graine :

Ces graines ont des propriétés toxiques, La toxine : la ricine est capable d’inhiber la synthèse des protéines en attaquant l'ARN des ribosomes,
entraînant la mort cellulaire.
Elle est présente à une concentration variant de 1 à 10 % dans la graine de ricin.
Il est réalisé une coupe du péricarpe qui enveloppe la graine :
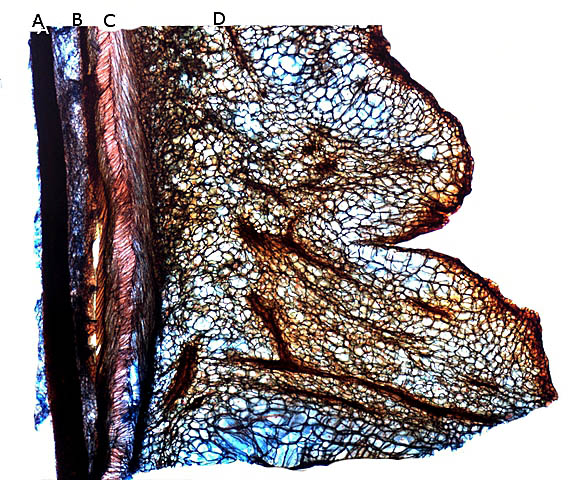
A -- Téguments de la graine.
B -- L'endocarpe est la couche la plus interne du péricarpe. Il est généralement mince et adhère à la graine
C – Le mésocarpe est la couche intermédiaire du péricarpe. Il est généralement plus épais que l'épicarpe
D – L'épicarpe est la couche la plus externe du péricarpe. C’est cette couche qui donne cet aspect spectaculaire à ce fruit grâce au développement d’ importantes ornementations .
La graine :

Les graines seront évacuées par des déhiscences latérales qui au mois d’ Aout ne sont pas encore visibles -
Classification classique
Règne Plantae
Classe Magnoliopsida
Ordre Euphorbiales
Famille Euphorbiaceae
Genre Ricinus
Nom binominal
Ricinus communis
L., 1753
Pour en savoir plus :
https://fr.wikipedia.org/wiki/Ricin_commun
La totalité de la plante semble toxique en raison de la présence d'une lectine glycoprotéique : la ricine. La concentration en ricine est maximale dans les graines qui renferment par ailleurs des protéines, de l'eau et des lipides. Ces graines sont riches en une huile qui doit ses propriétés purgatives à la présence de l'acide ricinoléique. qui altère la muqueuse intestinale et provoque des pertes importantes en eau et en électrolytes (sels minéraux), d'où son action purgative intense et irritante. La ricine, protéine présente dans la plante et les graines, est une toxine . . Le passage à l'autoclave de la farine pendant 15 minutes à 125 °C détruit la ricine.
On considère que trois graines peuvent être fatales à un enfant, quatre graines peuvent déterminer une intoxication sérieuse chez l'adulte et six à huit graines pourront lui être fatales. Les pigeons ramiers sont également sensibles aux graines de ricin et de nombreux cas d'intoxication ont été constatés dans les villes qui en utilisaient comme plante ornementale.
La production des graines de ricin dépasse les 100 millions de tonnes par an ( Production surtout en Inde - en Chine et au Brésil )
On l'utilise depuis peu comme source de biocarburant. L'huile est également utilisée comme lubrifiant dans les moteurs de voitures de course et de modèles réduits.
Il entre dans la composition d'une matière plastique (non-biodégradable) nommée Rilsan.
Dominique.
-
Samare
A cette période de l’ année certains arbres de nos forêts se couvrent de gros bouquets de formations ailées - Il s’agit de la période de fructification –Les samares sont un moyen utilisé par certains arbres afin de disséminer leurs graines - Les Erables - les Frênes sont les plus fréquents de la région - (Les Ormes ont disparu il y a quelques années ).
Définition :En botanique, la samare est un fruit sec indéhiscent, c'est-à-dire un akène (contenant une seule graine), muni d'une excroissance en forme d'aile membraneuse, formée par le péricarpe.
L’ Érable sycomore:

La samare double ou disamare (qui dérive d'un ovaire à deux carpelles) est le fruit des érables (Acéracées). Cette photo montre que le Gynécée de la fleur peut parfois être constitué de 3 carpelles et par conséquent donner naissance à une trisamare.
La samare est donc constituée de deux parties - une aile qui présente un bord d’ attaque et une asymétrie entre les deux faces
- un fruit arrondi situé à ‘extrémité supérieure de la samare ( on verra plus tard les raisons aérodynamiques de cette position )
Coupe d’une samare du sycomore:

Cette coupe met en évidence une symétrie de structure des faces supérieure et inférieure de la Samare
Le fruit du Sycomore est porté à l’ extrémité de la samare.
Ce fruit est constitué du péricarpe et de la graine
Le péricarpe :
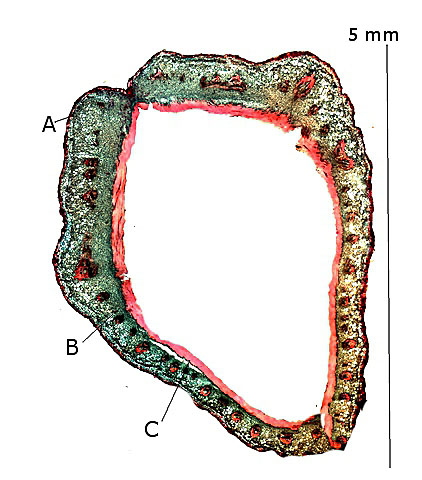
Le péricarpe est divisé en trois couch,
A – L’ épicarpe ( ou exocarpe ) qui est une couche mince et durcie ( non ligneuse) .
B -- Le mésocarpe ( qui est chez beaucoup de fruits la partie succulente ici il existe simplement plusieurs couches de cellules avec de nombreux axes vasculaires.
C – L’ endocarpe de consistance variable suivant les fruits mais qui ici est ligneuse.
------ L’ épicarpe (la peau) est habituellement une mince couche durcie de couleur bien que généralement pas ligneuse; cette couche est encore nommée l'exocarpe;
------ Le mésocarpe (la chair) est habituellement construit par de nombreuses grandes cellules et constitu une partie succulente du fruit;
------ L'endocarpe peut avoir une consistance similaire à celle de mésocarpe ou durcir beaucoup.
Certains types de fruits ont leur péricarpe fusionné avec le tégument de la graine en une seule couche appelée caryopse (le grain). Les plantes concernées sont des herbacées, essentiellement des céréales comme : le blé, le maïs et le riz, et la plupart des céréales cultivées dans le monde.
La graine:
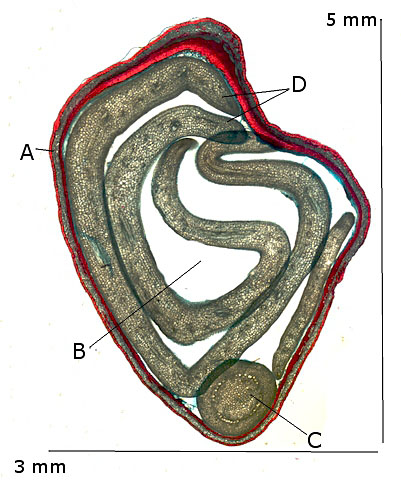
La graine
- A – Le Tégument : ce tissu constitue le tissu protecteur il est constitué d’une couche de plusieurs cellules qui entoure complétement l’ embryon et ses réserves - .Il existe cependant un petit orifice le micropyle , où a pénétré le tube pollinique pour la fécondation .Chez l’ Erable sycomore le tégument n’ adhère pas au péricarpe..
- B -- Albumen : Dans cette graine il n’ y a pas d’ albumen ; elle est dite Exalbuminée ,dans ce cas les réserves sont stockées directement dans les cotylédons.
. . C – L’embryon : Dans notre observation l’embryon n’est pas développé et se limite à une masse cellulaire non différenciée.
D – Les deux cotylédons qui assureront la photosynthèse dès leur apparition à la lumière solaire.
Histologiquement existe -il une différence d’organisation entre l’ aile d’une samare et une feuille ?
Il est réalisé une coupe des deux éléments .
La feuille :

On retrouve chez la feuille une asymétrie entre les faces exposées à la lumière et les faces inferieures exposées à l’ ombre
A – Cuticule
B -Couche de cellules palissadiques
C - Couche dite lacunaire ( liée aux échanges gazeux avec l’ extérieur grâce aux stomates )
D - Axe vasculaire contenu dans une nervure fait de la reunion d’un grand nombre de vaisseaux
La samare :
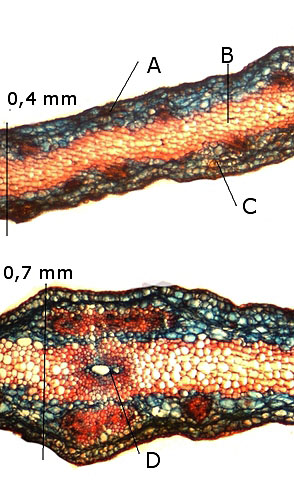
A – Couche de cellules épithéliales.
B - Parenchyme médullaire.
C - Couches de cellules epitheliales identiques à celles de l’ autre face de la samare.
Cette organisation s’explique par le fait que l’ aile d’une samare n’ est fait que grâce à l’ extension du péricarpe du fruit .
D - 1 seul vaisseau au centre de la nervure de la samare.
Le frêne .

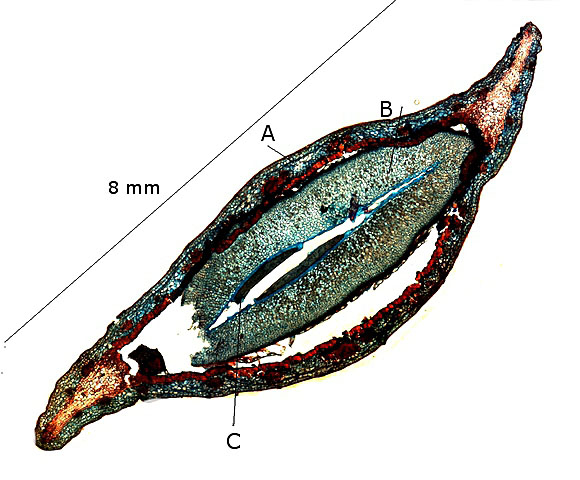
A -- Péricarpe.
B – Cotylédons . .
C --L’ embryon n’ est pas visible la coupe ayant été endommagée lors de sa réalisation
Le Charme
Selon le site https://www.zoom-nature.fr/identifier-les-arbres-a-samares/
On peut regrouper les différentes essences à samares en groupes informels (G) selon la forme des samares.
Samares en forme de grosses pièces avec une aile tout autour et la graine au milieu : G1) Ormes (Ulmus) et orme de Samarie (Ptelea). du tulipier de Virginie, de l'ailante ou de certaines Légumineuses (Tipuana tipu);
Samares en forme de langue allongée : G2) Frênes (Fraxinus) et Ailante (Ailanthus)
Samares disposées par deux : G3) Érables (Acer)
Samares différentes : voir G4) Le Charme fait partie de ces samares moins caractéristiques - le tilleul argenté aussi -



Cette samare permet de mettre en évidence l’ origine de l’ aile :
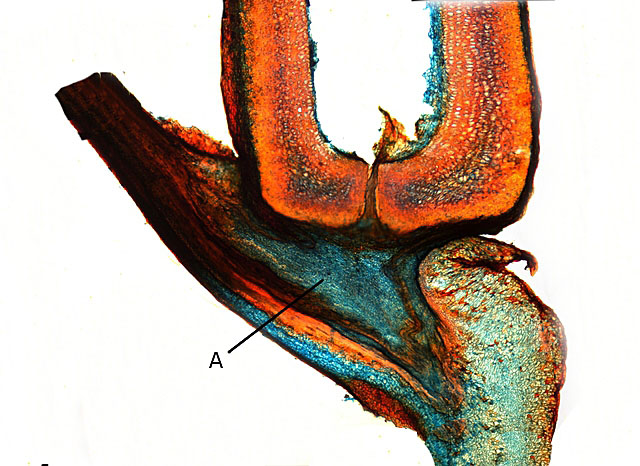
A - zone d’ hypertrophie du péricarpe dont l’extension va conduire à la mise en place des ailes
Commentaire :
Pour comprendre comment une samare vole il faut se reporter à l’ article suivant : https://les-samares.webnode.fr/forces-et-portance/ ++++
L’ étude porte sur la samare de l’ Erable .
Il y est analysé pourquoi l’ aile de la samare est efficace sans avoir toutes les caractéristiques de l’ aile d’ avion ou de la pale de l’ hélicoptère
Ainsi, l'aile de la samare, de par sa forme qui lui permet de conserver sa vitesse semble avoir la possibilité de générer une force de portance.
L'angle d'incidence élevé adopté par la samare lors de sa chute pourrait générer un différentiel de pression entre l'extrados et l'intrados, et donc générer une force de portance.
Cet angle d'incidence élevé, ainsi que le profil de l'aile de la samare, plus plat que celui d'une aile classique ou d'une pale d'hélicoptère conduit à la formation d'un mini vortex sur l'extrados de l'aile.
En conclusion
https://fr.wikipedia.org/wiki/An%C3%A9mochorie
L’anémochorie est la dispersion par le vent des diaspores. Il s'agit d'un des trois types de dissémination assurée par un agent extérieur (allochorie).
L'anémogéochorie est une sous-catégorie d'anémochorie qui désigne les virevoltants dont la dispersion se fait près du sol, mais dont le vent reste l'agent dispersant.
La forme de l'aile, qui a pour fonction d'aider à la dissémination du fruit, est variable : allongée chez le frêne, dissymétrique chez l'érable, circulaire enveloppant l'akène chez l'orme en trois parties chez le Charme .
Dominique.
-
Bonjour Patrick
Ton éclaircissement est au top : ni trop - ni trop peu.
Amicalement
Dominique.
-
Bonsoir Patrick
Parmi les éclaircissants on trouve
Pour les insectes : Solution de KOH 10% 24 h puis 2 heure dans l’acide lactique à 25 % ( il est conseillé de surveiller le niveau d’ éclaircissement ).
Ou solution aqueuse acide lactique à 60% en chauffant un peu on réduit les temps d’éclaircissement ..
L’eau Oxygénée à 30 volumes décolore la mélanine en 24 à 48 h .
Le permanganate de potasse à 10% est actif après 2 à 3 h
Pour le phénol il est utilisé comme désinfectant et antiseptique, ainsi que dans des préparations médicales telles que les bains de bouche et les bonbons pour maux de gorge.
on l' utilise dans l' industrie alimentaire pour désinfecter les tuyauteries en inox ( suivi d' un bon rinçage ) tout est une question de dosage .
Amicalement
Dominique
-
Bonsoir Jean- Marie
Ton travail est tout à faire remarquable - pouvoir le découvrir est un réel plaisir
Amicalement
Dominique
-
Bonsoir Patrick
Merci et à bientôt pour de belles photos.
Amicalement
Dominique
-
Bonsoir Jean -Claude
Ta proposition est très agréable ,mais en dehors des professionnelles , je ne sais pas si quelqu'un va avoir assez d' orties dans sa propriété pour obtenir une quantité tissable.
Amicalement
Dominique
-
Feuille de maïs adaptation à la sécheresse
En période de sécheresse les feuilles de certaines plantes vont se déformer et se ratatiner.
C’est le cas de la feuille de maïs dont la transformation est bien visible en période de manque d’eau.

Après une période de sécheresse :

Une coupe de la feuille explique ce phénomène de repliement :
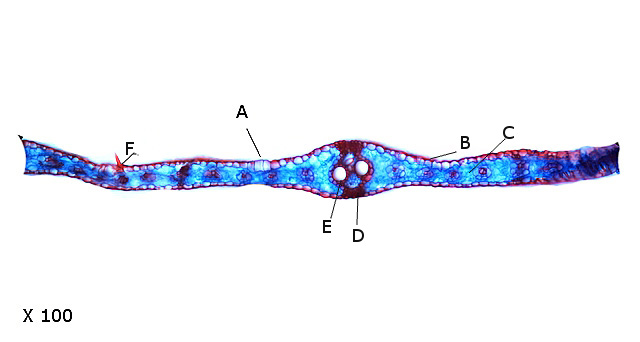
A – Cellules bulliformes ou motrices.
B – Cellules épidermiques.
C—Parenchyme (ici il n’existe pas de parenchyme palissadique.)
D—Nervure (Axe vasculaire).
E—Vaisseaux du Xylème.
F-- Trichome.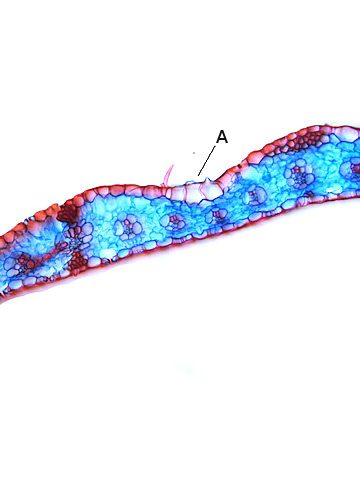
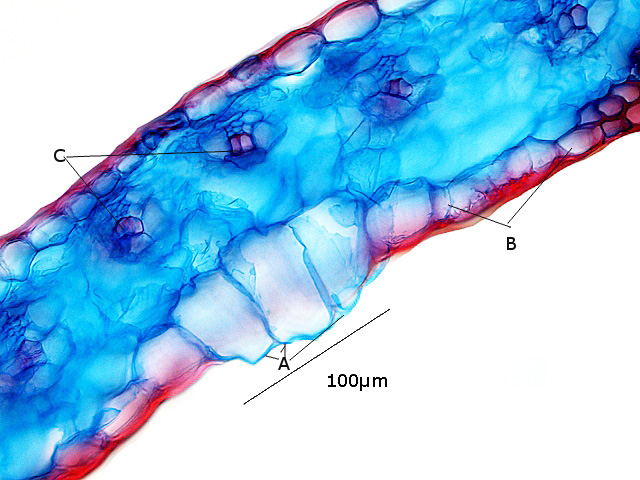
A – Cellules bulliformes
B – Cellules épidermiques couverte de la cuticule ( melange de cutine et de cires ).
C – Nervilles ( nervure de troisième rang )
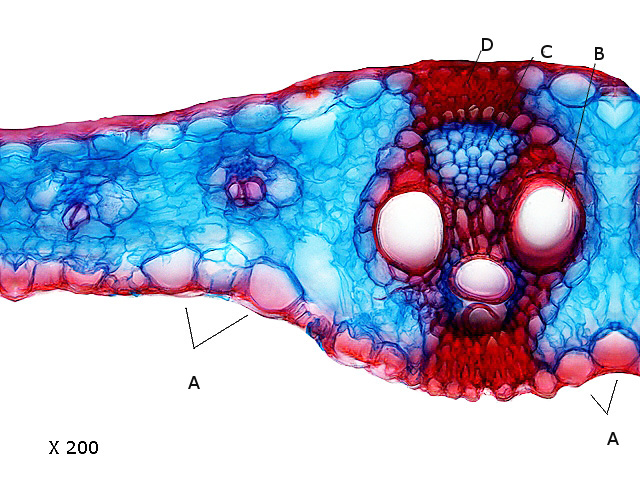
A -- Cellules bulliformes.
B -- Xylème.
C -- Phloème.
D - -Sclérenchyme. ( qui assure la rigidité de la nervure )
L’épiderme des graminées est composé de divers types de cellules.
La plupart des cellules épidermiques sont étroites et allongées - certaines cependant sont particulièrement grandes : ce sont les cellules bulliformes ou cellules motrices.
Elles sont disposées en rangs longitudinaux sur la face inferieure des feuilles de maïs.
En perdant leur eau elles pourraient être responsables du repliement ou de l’ enroulement ( dans la cas de la feuille de maïs) en relation avec les modifications du potentiel hydrique .En cas de manque d’ eau ces cellules deviennent flasques et la feuille se replie ( si les cellules bulliformes sont situées de part et d’autre de la nervure principale) ou s’enroule (si les cellules bulliformes sont reparties plus ou moins régulièrement sur la surface du limbe de la feuille ) - Inversement si la charge en eau augmente -
Sur la préparation on constate que ces groupes cellulaires sont positionnés très près des nervures qui servent d’ axe de rotation .
Référence
Biologie végétale Raven Evert Eichhorn éditeur deBoeck page 599
https://www.arvalis.fr/infos-techniques/ensiler-ou-non-un-diagnostic-prealable-simpose
Dominique.
-
Feuille le système vasculaire
Feuille de Liriodendron tulipifera ( Tulipier de Virginie )
:
Comment s’organisent les nervures et comment cela fonctionne –t –il ?
Pour essayer de comprendre le système une feuille de Tulipier de virginie va être utilisée - ( il y plusieurs Tulipiers dans le jardin botanique voisin dont les feuilles se confondent avec celle de l’ érable sycomore ).

Si on place cette feuille dans un bain d’eau oxygénée 2 jours suivi d’un bain dans l’ eau de javel 4 jours le décoloration est totale et le système des nervures devient bien apparent.

La nervation de la feuille de tulipier est dite réticulée ( pour les monocotylédones on parle de nervures paralléles )
1--nervure principale médiane.
2--nervures principales latérales.
3-- nervures de deuxième ordre. ou nervioles
4—nervures de troisième ordre ou nervilles.
La feuille reçoit ses nutriments par le pétiole qui aussi envoie vers l’arbre les molécules fabriquées par la photosynthése dans la feuille.
Le pétiole:
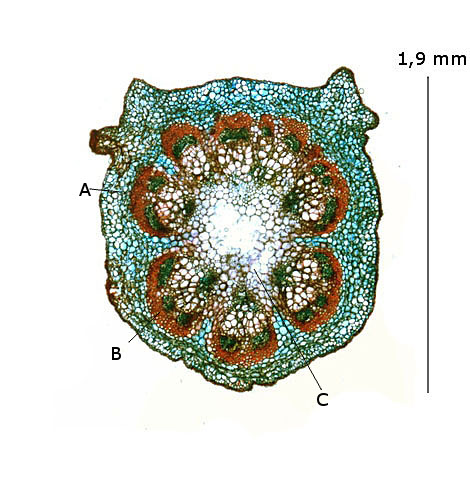
A –Parenchyme parietal.
B – Axes vasculaires
C –Parenchyme médullaire
Quel est l’aspect de chacune de ces nervures
Nervure principale médiane :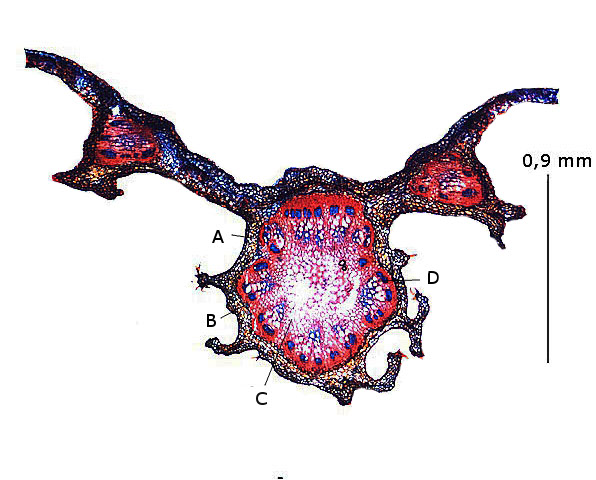
A – Parenchyme pariétal.
B -- Péricycle.
C – Parenchyme médullaire.
D --Axe vasculaire
Nervure de deuxième rang.ou nerviole.
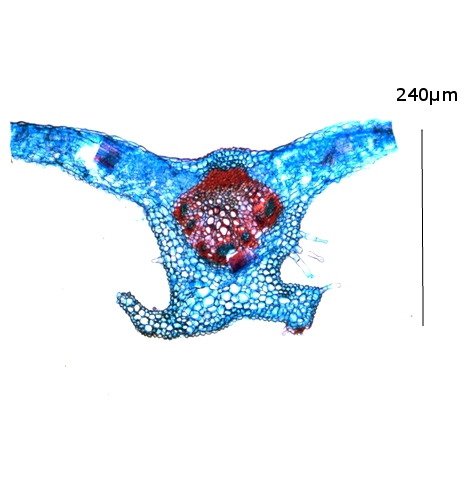
Nervure de troisième rang ou nerville:
A - Cellules de la gaine fasciculaire (voir plus loin).
B - Vaisseaux de Phloème
C - Vaisseaux de Xylème .
Dans les feuilles le plus souvent les vaisseaux de phloème sont proches de la face supérieure tandis que les vaisseaux de Xylème sont plus proches de la face inferieure
Les petites zones du mésophiles (parenchyme chlorophyllien) délimitées par ces nervures ( sans chlorophylle ) de troisième rang sont appelées aréoles .
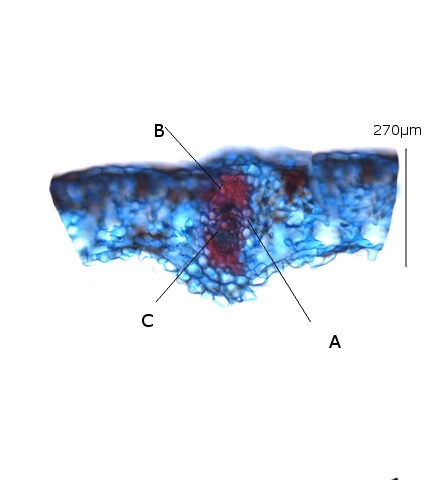
Les aréoles :
A – la gaine fasciculaire : située entre les vaisseaux et le parenchyme. (ces cellules n’ont pas de chlorophylle).
B – les trachéides
Dans ces aréoles il est mis en évidence des lignes sans chlorophylle ; ce sont les trachéides qui forment la partie terminale des vaisseaux du Xylème (apportent l’ eau à l’ épiderme ) . Il existe aussi des fibres de phloème qui vont drainer les produits fabriqués pas les cellules du mésophile. Ce transfert se fait toujours à travers la gaine fasciculaire. (A).
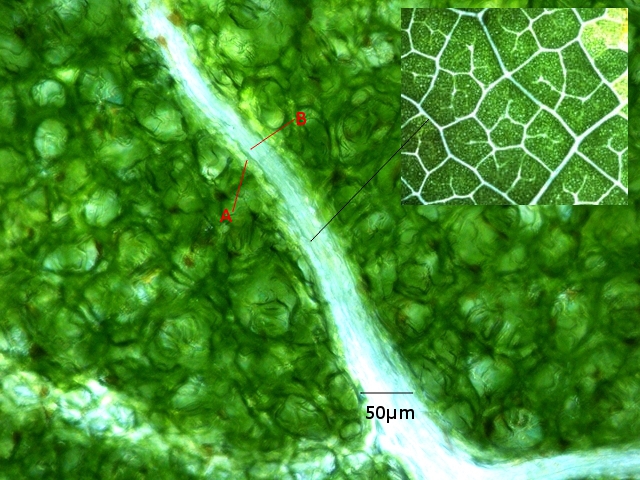
Aspect de la partie terminale du système de nervation :
A-- Cellules de la gaine fasciculaire.
B –Trachéides situées au centre de la gaine.
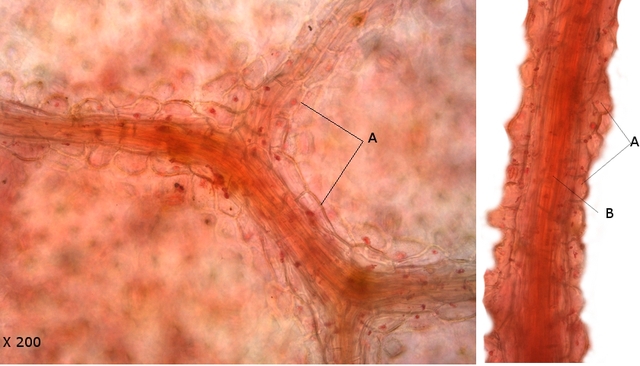
Vue longitudinale externe de l’organisation des trachéides :
A --Les trachéides sont des conduits permettant le transport de la sève brute (composée d'eau et de sels minéraux) chez les plantes vasculaires. Elles sont composées d'une suite de cellules de forme allongée. Lors de la formation des trachéides, ces cellules meurent, sont vidées de leur contenu et sont fusionnées
verticalement pour former un conduit creux. ; Elles possèdent aussi un rôle dans le soutien global de la plante.
B – cellules de la gaine fasciculaire.
Conclusion :
Ces images mettent en évidence la structure terminale des vaisseaux :
Les vaisseaux ne sont jamais en contact direct avec les cellules du mésophile .Ces vaisseaux sont toujours entourés par une ou deux couches cellulaires la gaine fasciculaire qui va jusqu‘au bout des tissus conducteurs ;donc mésophile et tissus conducteurs ne sont jamais en contact direct.
Tout ce qui pénètre et sort de ces tissus passe obligatoirement au travers de cette gaine fasciculaire.
Il ne faut pas oublier que les vaisseaux de lignine sont perméables à l’’eau ( vaisseaux criblés par des pores ), donc les liquides peuvent passer au travers de la paroi des vaisseaux puis ensuite passer au travers de la gaine ; tel est l’ organisation du système d’ échanges au sein de ce laboratoire de chimie qu’ est une feuille .
*********************************
Aspect d’une coupe de la feuille du Tulipier de Virginie
A – l’épiderme et sa cuticule d’une structure compacte qui augmente la résistance physique de la feuille.
B – les cellules palissadiques - ces cellules ne sont pas aussi bien rangées que dans les feuilles de plantes annuelles ( on reviendra sur cet aspect plus tard).
C – parenchyme spongieux cette zone est riche en espaces aériens qui s’ouvrent sur l’extérieur par les stomates.
D - stomate.
E – espace intercellulaire.
Référence
Biologie végétale de Raven – Evert –Eichhorn éditeur deBoeck page 583 et suivantes
Dominique.
***************************************************
Pour en savoir plus : Déterminisme des mouvements de la sève.
La sève brute :
La sève brute, qui contient de l'eau et des minéraux absorbés par les racines, est principalement transportée dans les vaisseaux conducteurs appelés xylème. Ces vaisseaux se trouvent dans les parties centrales de la plante, notamment dans les tiges et les racines.
Le mouvement ascendant de la sève brute repose sur plusieurs mécanismes :
Transpiration : Les plantes ont des ouvertures microscopiques appelées stomates, présentes principalement sur les feuilles. Lorsque les stomates s'ouvrent, l'eau s'évapore des cellules qui tapissent les espaces intercellulaires de la feuille et s'échappe dans l'atmosphère sous forme de vapeur d'eau. Ce processus est appelé transpiration.
Cohésion : L'eau est une molécule polaire, ce qui signifie qu'elle est attirée par d'autres molécules d'eau grâce aux forces de cohésion. Ces forces de cohésion permettent à l'eau de former une colonne continue à l'intérieur des vaisseaux du xylème.
Tension : Lorsque l'eau s'évapore des feuilles, cela crée une tension dans la colonne d'eau présente dans les vaisseaux du xylème. Cette tension tire l'eau vers le haut, contre la force de gravité, à travers les vaisseaux.
Capillarité : Les vaisseaux du xylème sont étroits et minces, ce qui permet à l'eau de monter par capillarité. La capillarité est la tendance de l'eau à monter dans de petits espaces, tels que des tubes étroits, en raison des forces d'adhésion entre l'eau et les parois des vaisseaux.
La sève élaborée :
Le mouvement de la sève élaborée, qui contient principalement des sucres et autres substances organiques produites par la photosynthèse, est principalement dû à un processus appelé pression de turgescence ou pression osmotique. Ce processus repose sur la différence de concentration en solutés entre les cellules du phloème et les cellules environnantes.
Lorsque la plante produit des substances organiques, telles que des sucres, dans les cellules du phloème, cela crée une concentration plus élevée de solutés à l'intérieur des cellules du phloème par rapport aux cellules environnantes. En conséquence, l'eau circule par osmose des cellules environnantes vers les cellules du phloème, créant une pression de turgescence.
La pression de turgescence pousse la sève élaborée vers les régions de plus faible pression, ce qui permet à la sève de se déplacer dans les vaisseaux du phloème et d'être distribuée vers les parties de la plante qui en ont besoin, telles que les racines, les bourgeons, les fleurs et les fruits.
De plus, la photosynthèse dans les feuilles crée une source supplémentaire de sucres, ce qui entraîne une augmentation de la pression osmotique dans les cellules du phloème à proximité. Cela contribue également à la poussée de la sève élaborée dans les vaisseaux du phloème.
Il est important de noter que le mouvement de la sève élaborée dans le phloème est un processus actif qui nécessite l'énergie métabolique de la plante. Les cellules du phloème, appelées cellules compagnes, jouent un rôle crucial en utilisant de l'énergie pour transporter activement les substances organiques dans les vaisseaux du phloème et maintenir le mouvement de la sève élaborée.
-
Chardon évolution de la tige en fonction de sa position.
Dans le jardin il y a toujours eu des chardons - ce n’ est pas très ennuyeux car on peut les arracher facilement - mais les graines produites par un seul pied sont si nombreuses qu’ un seul pied est capable de réensemencer tout le jardin.
Dans un coin un peu plus recule j’ ai marché sur une tige de chardon sans le remarquer Quelques jours après la tige en partie écrasée à modifier son organisation .
La tige a développé plusieurs nouvelles pousses et à la place d’ un seule tige 3 sont apparues dans notre observation - mais pas encore de racine - il faut remarquer qu’en ce mois de juin il ne pleut plus en Normandie ( ou si peu ) depuis plusieurs semaines ce qui rend la terre extrêmement dure .


Aspect d’une tige normale non horizontalisée au même niveau que la tige précédente.

Coupe d’une tige habituelle de chardon
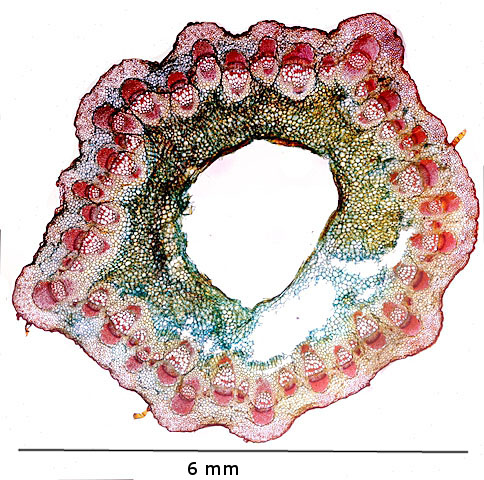
Sur cette photo d’une coupe de la tige du chardon le nombre d’axes cribro-vasculaires est très important en comparaison avec les autres plantes.
L’organisation d’un axe normal.
Par définition Un faisceau cribro-vasculaire,( appelé aussi faisceau conducteur, vaisseau conducteur ou faisceau libéro-ligneux) est une partie du système circulatoire d'une plante vasculaire . l
L’origine de ce système est le méristème latéral ( 2 ) ( encore appelé méristème secondaire ou cambial ) ;
Les cellules méristématiques indifférenciées acquièrent alors une structure et une fonction spécifique.
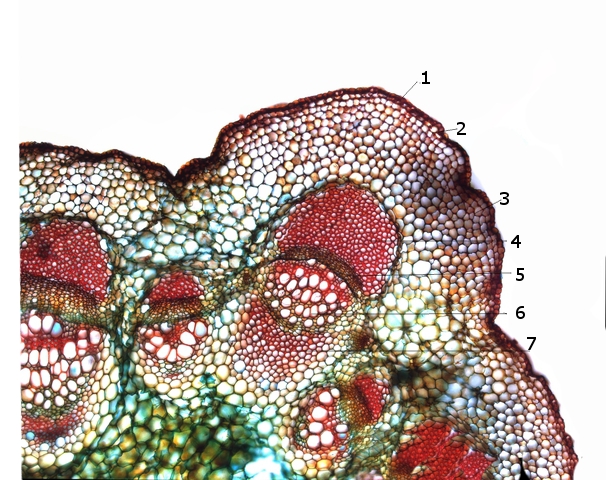
1 – Cuticule .
2 – Epiderme.
3 – Parenchyme cortical et collenchyme (ce dernier soutient et renforce ces régions en leur donnant une structure flexible. Il est responsable de la résistance mécanique des tissus végétaux, ) .
4 – Phloéme .
5 – Cambium Les cellules cambiales sont généralement disposées en couches concentriques,( formant des anneaux de croissance). Lorsque les cellules cambiales se divisent, certaines d'entre elles se différencient en cellules xylémiques, qui sont responsables du transport de l'eau et des minéraux, tandis que d'autres se différencient en cellules phloémiques, qui assurent le transport des nutriments organiques. Ce processus de division et de différenciation des cellules cambiales permet l'expansion du xylème et du phloème, entraînant ainsi la croissance en épaisseur de la plante.
6 – Xyléme.
7 –Protoxyléme ( proto : formé avant ) ( meta : formé aprés) et gaine sclerenchymateuse (le sclérenchyme est un tissu végétal mort, composé de cellules avec des parois cellulaires épaissies et lignifiées, qui jouent un rôle majeur dans le renforcement et le soutien structurel des plantes.)
Or c’est le phénomène de développement de l’ activité de ces méristèmes secondaires , non plus dans le sens - centre -périphérie - mais dans un sens perpendiculaire qui se met en marche dans notre observation et qui est à l’ origine des nouvelles tiges apparues sur la tige couchée du chardon .
Coupe de la tige au niveau de l’apparition des nouvelles tiges:
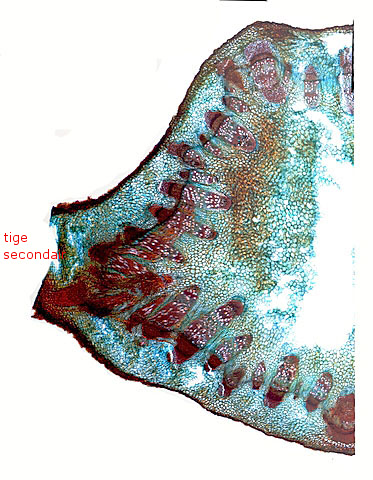

En A la photo met en évidence l’apparition (dans un plan différent de celui de la croissance normale de la tige) d’un tissu nouveau qui va combler l’ espace entre deux axes vasculaires adjacents et se poursuivre à travers les tissus corticaux pour permettre le développement d’une nouvelle tige non prévue dans le plan initial de la plante.
Aspect à l’union tige mère et de la nouvelle tige
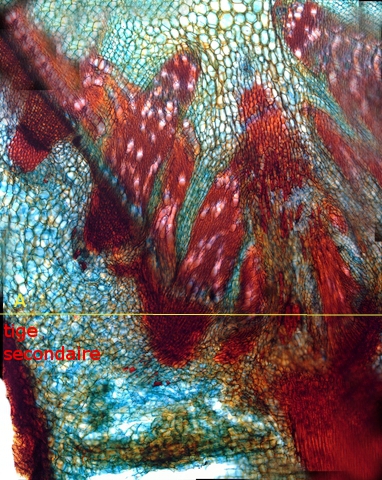
La ligne jaune ( A ) marque la séparation entre les limites de la tige mère et de la nouvelle tige .Le système cribro-vasculaire est apparemment désorganisé dans sa structure mais il peut désormais assurer son rôle de nourrisseur , de drainage de la nouvelle structure et de croissance en épaisseur .
Conclusion :
On constate que les méristèmes latéraux responsables de la croissance des tiges des plantes en épaisseur vont s’ activer à la suite de la modification de la position de cette tige par rapport à l’ horizontale.
Les coupes réalisées montrent que seulement quelques méristèmes latéraux vont modifier et accélérer leur prolifération cellulaire .
Ce sont les méristèmes situés dans le plant perpendiculaire à la nouvelle direction de la tige.
La plante a donc perçu la modification d’ orientation et a déclenché la processus de multiplication uniquement dans une direction donnée ).
Le méristème apical ( celui situé au sommet de la nouvelle tige ) va réagir quant à lui à l’ orientation de la lumière et de ce fait permettre à la nouvelle tige de retrouver la verticalité.Ce processus est déclenché sur plusieurs zones de la tige couchée ( dans le cas présent environ tous les 5 centimètres ) distance nécessaire pour que les nouvelles tiges ne se gênent pas .
Mais comment ces modifications sont-elles perçues par la plante ? qui prend la décision de ces modifications ? Qui coordonne le planning de cette nouvelle restructuration ? La réponse moderne est l’ADN et la participation des processus d’épigénétique déclencheurs des réactions biochimiques.
Mais tout cela reste quand même merveilleux pour les ’observateurs que nous sommes.
***********************************************
Pourquoi Cribro – vasculaire ?
Dans le contexte de la botanique, l'expression "axe cribro-vasculaire" fait référence à une caractéristique structurelle spécifique d'une tige végétale.
L'axe cribro-vasculaire est une région particulière de la tige où se trouvent les tissus conducteurs primaires, à savoir le xylème et le phloème. Le xylème est responsable du transport de l'eau et des minéraux depuis les racines vers les parties aériennes de la plante, tandis que le phloème assure le transport des nutriments, tels que les glucides, dans toutes les parties de la plante.
La caractéristique distinctive de l'axe cribro-vasculaire réside dans la présence de cellules criblées dans le phloème. Ces cellules criblées, également appelées cellules criblées du phloème, sont interconnectées par des régions criblées, qui sont des zones perforées permettant le passage des nutriments à travers le phloème.
Référence
Manipulation de botanique Duno éditeur de L Genevés
( Avec l’ aide de Chatgpt )
Dominique
-
Blé épi et épillet ( le blé en fleur )
Le genre Triticum appartient à la famille des Graminées, ou Poacées. Parmi toutes les espèces de blé cultivées, le Triticum aestivum, ou blé commun, est la plus importante sur le plan économique.
Depuis quelques jours le blé est en fleur - La fleur est bien discrète et nécessite pour être vue un examen attentif de l épi.

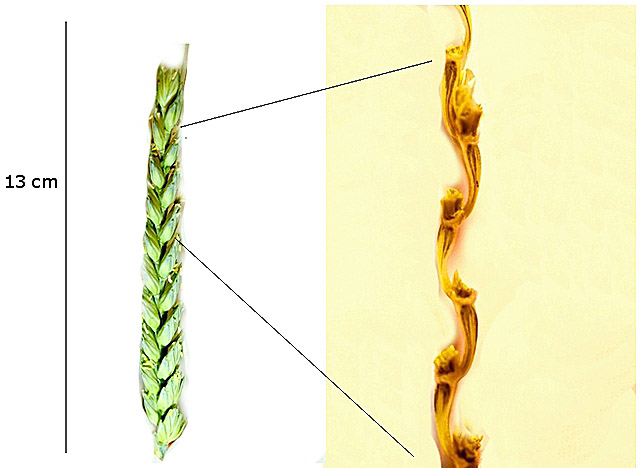
L’épi de blé est une inflorescence constituée d’unités appelées épillets.
La fixation de ces épillets se fait directement sur l’axe de l’inflorescence. Le blé a des épillets possédant un pédoncule très court en position alternée comme le montre la photo de droite.

L’épi de blé présente -- deux faces ( antérieure et postérieure) qui sont formées par la superposition des épillets( vus de face).
-- deux faces latérales.
L’épillet est une inflorescence secondaire :
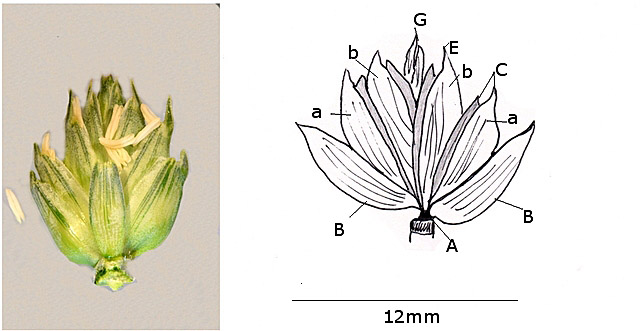
L’épillet : sa constitution :
A – Le pédoncule très court.
B -- Les glumes au nombre de 2 sont des pièces foliacées en position alternée .Leur rôle est de protéger la base de l’épillet .
C -- Les glumelles qui sont aussi en position alternée.
D - Pour chaque fleur il y a 2 glumelles la glumelle inférieure ou lemme.
la glumelle supérieure ou paleole .
Leur rôle est de protéger la graine.
E -- La glumelle inférieure est parfois porteuse d’une arête qui dans certaines variétés peut être très longue.
F - L’épillet est ici constitué de 4 fleurs (toutes ne sont pas fertiles).
a- les fleurs primaires qui seront les premières à ouvrir
b- les fleurs secondaires qui s’ouvriront ensuite
G -- Le sommet de l’épillet est formé de deux fleurs sans ovaire ni étamines.
La fleur :

Les deux glumelles ( supérieure et inférieure ) forment une cavité fermée où se développe la fleur.
Ce n’est qu’au moment de la pollinisation que les anthères vont émerger vers l’extérieur.
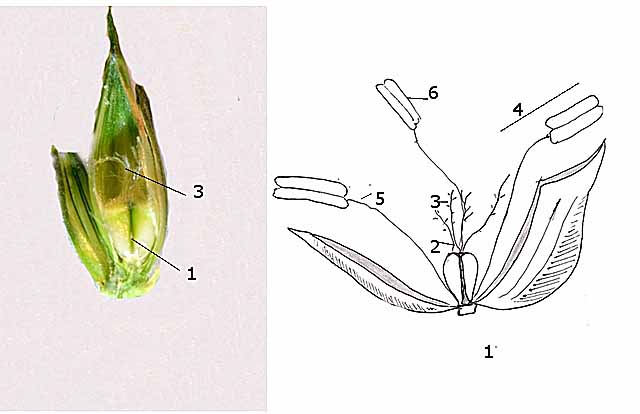
La fleur est formée par
1 – Ovaire
2 -- Styles ( 2 styles ramifiés )
3 -- Stigmates
4 -- Étamines chez les poacées il y a 3 étamines.
5 - Filet
6 -- Anthères
L’ anthère - chez le blé il y a une seule anthère par étamine ( chaque anthère a deux loges ).
Une anthère est la partie terminale de l'étamine, organe mâle de la fleur, qui produit et renferme le pollen.
L'anthère est fixée au filet de l'étamine par son milieu (anthère médifixe ou dorsifixe).
Elle est composée de deux thèques possédant chacun deux sacs polliniques avant maturité, nombre constant chez les Angiospermes. Ceux-ci fusionnent à maturité pour donner deux loges (anthère biloculaire .Elles s'ouvrent, généralement par déhiscence, pour répandre les grains de pollen à l'extérieur.
Le pollen :
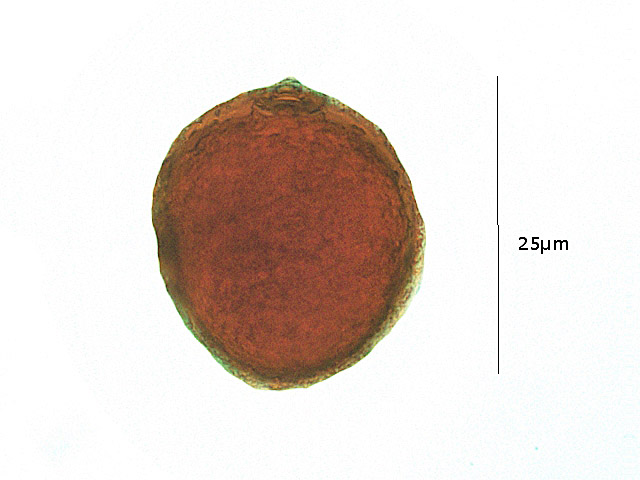
Au sommet du grain de pollen faisant saillie un peripore - qui permettra l'émergence du tube pollinique
La taille d'un grain de pollen de blé peut varier légèrement en fonction de différentes espèces de blé. En général, les grains de pollen de blé sont assez petits. Ils ont une taille moyenne d'environ 25 à 30 micromètres, Cette taille est en fait variable suivant des facteurs externes ..
.. L’exine ( couche externe) a une ornementation rugueuse .Au sommet fait saillie un pore germinatif unique ( monocotylédone ).
Le grain de pollen est le gamétophyte mâle, il produit et contient les gamètes mâles haploïdes (n1), et permet leur déplacement.
Le grain de pollen mature est constitué habituellement : de deux ou trois cellules non cloisonnées. Il comporte deux noyaux haploïdes : le plus gros est le noyau végétatif (qui donnera le tube pollinique), l'autre le noyau génératif ou reproducteur.
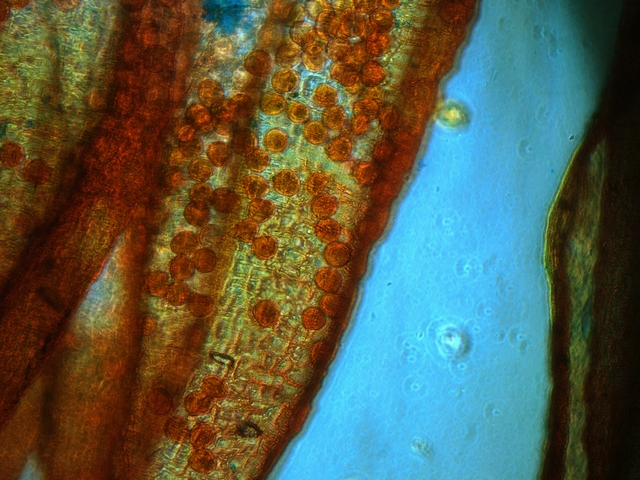
Sur cette photo en contraste de phase il est mis en évidence sur la paroi de l’ anthère un important réseau probablement vasculaire permettant la formation et le développement des grains de pollens.
Les stigmates :

Image des stigmates
Le pistil comprend un seul ovaire, un seul ovule et deux styles se terminant chacun par un stigmate plumeux et ramifié. Ces stigmates vont attirer les grains de pollens par électrostatisme . .
Le grain

Sa présentation a déjà été réalisée : https://forum.MikrOscOpia.com/topic/17981-grain-de-bl%C3%A9/
Ce fruit est un caryopse c’est à dire que le péricarpe et le tégument de la graine sont soudés - on parlera donc de graine
Le génie génétique permet aujourd'hui d'augmenter constamment le rendement du blé et la qualité du grain tout en diminuant la capacité du blé à survivre à l'état sauvage. D'ailleurs, le blé est cultivé depuis des siècles et on n'a relevé aucun cas où cette plante serait devenue envahissante au point d'être nuisible.
**********************************************************
Pour en savoir plus :
Le T. aestivum est hexaploïde (AABBDD), possédant en tout 42 chromosomes (2n = 42, soit six fois sept chromosomes). Le nombre haploïde de base est sept, puisque le nombre chromosomique des autres espèces de blé est également un multiple de ce nombre. Les cultivars de blé modernes sont soit tétraploïdes (blé dur, AABB), soit hexaploïdes (blé commun et blé club, AABBDD).
DeVries (1971) a constaté que les fleurs du blé demeurent ouvertes pendant 8 à 60 minutes, selon le génotype et les conditions écologiques. Au moment de la déhiscence des anthères, 5 à 7 % du pollen tombe sur le stigmate, 9 à 12 % reste dans l'anthère, et le reste est dispersé. Le pollen demeure ensuite viable pendant 15 à 30 minutes. Une fois libérés, les grains de pollen se dessèchent, et leur fixation aux ramifications du stigmate est assurée par une brève force électrostatique suivie de l'absorption d'eau par les grains de pollen à travers des ouvertures de la cuticule du stigmate (Heslop-Harrison, 1979). Ce processus permet en outre au tube pollinique de s'allonger, ce qui facilite la fécondation. Chez le blé, la période de réceptivité du stigmate dépend de la variété et des conditions écologiques mais se situe généralement entre 6 et 13 jours. La croissance du tube pollinique débute généralement une ou deux heures après la pollinisation, puis la fécondation survient 30 à 40 heures plus tard (deVries, 1971). Cependant, les graines de pollen peuvent germer peu de temps après leur contact avec la surface du stigmate et le blé peut être fécondé dans moins d'une heure (communication personnelle, George Fedak, 1999). Le premier épillet à fleurir se situe généralement dans le tiers médian de l'épi, habituellement près de la limite supérieure de ce tiers. La floraison progresse assez rapidement vers le sommet de l'épi et un peu plus lentement vers sa base. Les fleurs primaires de l'épillet s'ouvrent les premières, puis les secondaires, et ainsi de suite. Chez le blé, les étamines sont plus petites et produisent moins de pollen (1000 à 3800 grains de pollen par anthère, soit 450 000 par plant) que chez les autres graminées céréalières. Selon deVries (1971), ce nombre est de 4 millions chez le seigle (Secale cereale L.) et de 18 millions chez le maïs (Zea mays L.).
Référence
Dominique
-
Bonjour Jean -Luc
L 'article de W - demain parle d'un essai sur une grande surface agricole mais ne parle pas des techniques employées .Le problème avec l 'écologie c'est que ce n'est pas forcement la meilleur formule pour obtenir des résultats corrects sur le plan des rendements .L' ortie si elle pousse bien dans les terrains qui lui plaisent peut exiger des aménagements pour pousser dans les espaces ouverts .
Attendons les résultats des expérimentations en cours ..
Amicalement
Dominique.
-
L’ortie et les filatures
D’après le directeur du centre textile Lorrain Napoléon portait une veste tissée avec de la fibre d’ortie. La fibre d’ortie est donc la source d’une fibre que l’on peut tisser .
Son existence en tant que fibre textile remonte au néolithique. Ainsi, sur la momie Ötzi âgée d’environ 5300 ans découverte en 1991 sur un glacier des Alpes de l’Ötztal (entre l’Autriche et l’Italie), a été retrouvé un couteau dont le fourreau était fait de fibres d’ortie.
Au moyen âge la fibre d’ ortie était utilisée pour fabriquer des cordages.

Examen de la tige d’ortie qui sort du séchage :

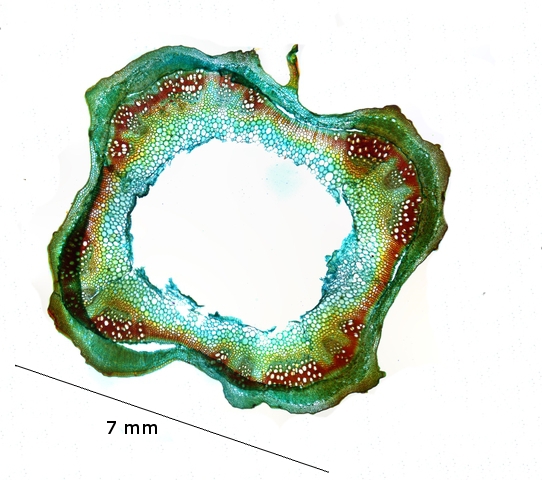
cette coupe d'une tige d' ortie montre que
La tige est de section quadrangulaire.
Les nervures de chaque angle sont formées par des gros cordons de collenchyme.
Examen en lumière polarisée :
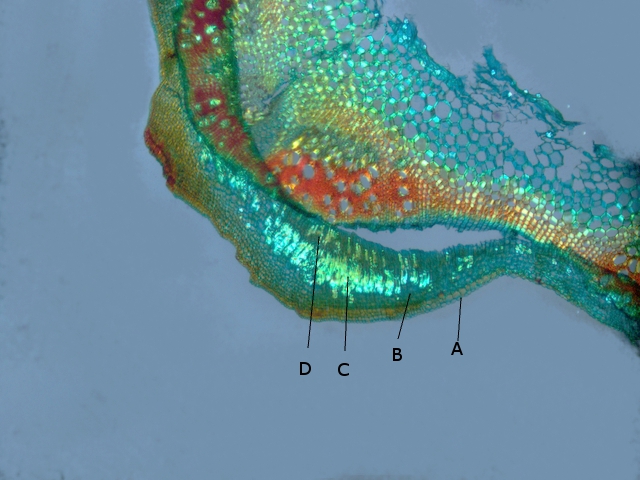
A—Epiderme
B –Collenchyme
C –Fibres
D –Parenchyme cortical
Coupe sur la longueur

A – Epiderme
B – Collenchyme
C – Fibre
D – Parenchyme cortical
E --Bois
F – Parenchyme médullaire
Examen en lumière polarisée:

La zone des fibres est bien apparente (A). *
La fibre se sépare assez facilement du bois

Examen en lumière polarisée
La fibre n’ est pas isolée .Elle se présente sous forme de faisceaux de fibre ( qui sont appelés fibre mais dans le cadre du tisserand seulement )
Exemple d’un faisceau : X 200 lumière polarisée:

Une fibre isolée X 200 lumière polarisée:

La fibre ne se présente jamais sous l’ aspect isolé - mais en faisceaux - comme pour le lin ce sont ces faisceaux qui sont appelés fibre dans l’ industrie.
Sa taille est d’un diamètre de :6 à 12 µm.
La longueur peut atteindre 8 cm
Cette fibre possède les propriétés mécaniques requises pour la filature ; soit une haute ténacité et une élongation de 2 à 4 % évitant toute rupture.
Il est intéressant de comparer l’aspect de la fibre de lin avec de la fibre d’ortie .
https://forum.MikrOscOpia.com/topic/12934-le-lin/?hl=%2Bfibre+%2Blin
Fibre de lin :

Une fibre de lin en lumiére polarisée :
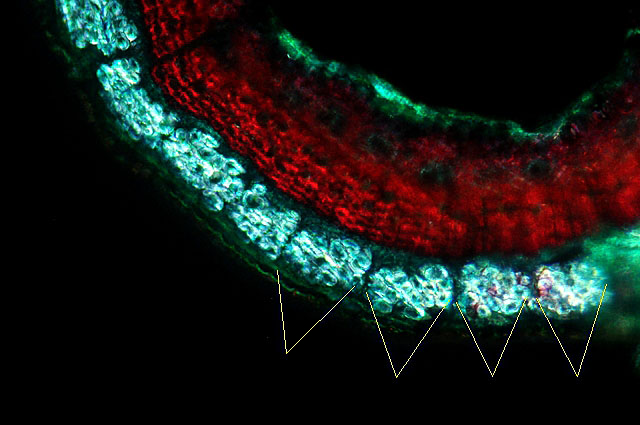
Chez le lin les fibres sont réparties en faisceaux tout autour de la tige.
Par contre pour l’ortie les fibres sont surtout localisées dans les nervures des angles de la tige .
La fibre d’ortie examen au X 400.
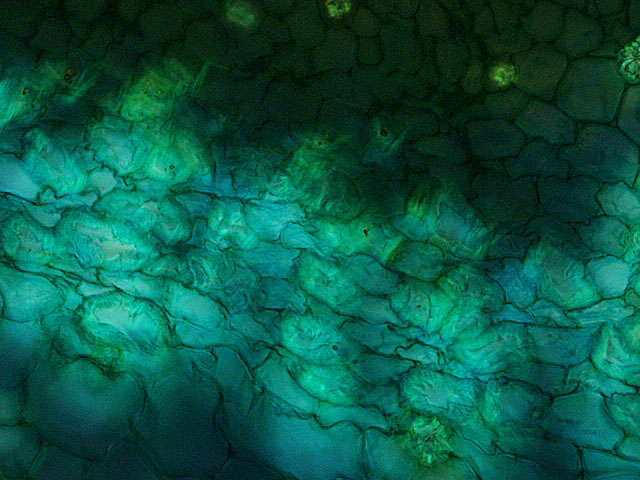
La différence entre les deux fibres
La fibre de lin est formée de cellules cylindriques - sa partie centrale( le lumen ) est bien distinguable .Les fibres sont de taille assez semblables.
La fibre d’ortie est formée de cellules aplaties- le lumen est peu visible. – leur organisation est plus désordonnée et surtout moins homogène dans leur taille .
Dans la présentation de la fibre de lin il est dit que la fibre peut se déformer ( donc que le tissu est froissable ) en raison de la présence de coudures ( les nœuds ) .On ne trouve pas ce type de formation sur la fibre de l' ortie .
**************************
Préparation de l’ortie pour l’industrie.
Cueillir l’ortie lorsqu’elle est arrivée à maturité.
Séparer la tige des feuilles.
Effectuer un rouissage pour détacher les fibres des déchets de bois difficiles à retirer.
Laisser sécher les fibres 2 jours environ.
Réaliser le battage des fibres pour casser le bois des tiges et commencer la séparation du bois.
Effectuer le teillage pour séparer les fibres de toutes les impuretés et ainsi obtenir une filasse brute.

La fibre d’ ortie sera filée afin d’ obtenir un fil solide , de grande taille et qui pourra être tissé ensuite. Ce fil sert à la fabrication entre autre de T-shirt - de Jeans et de très nombreux autres produits.
Cette fibre est aussi un excellent isolant qui peut être modelé en panneaux .
L’ortie se coupe 2 fois par an. Une fois qu’elle est implantée elle repousse tous les ans - elle demande peu d’ eau et pas d’ engrais ..
Son utilisation est donc beaucoup plus écologique que l’utilisation du coton dont la culture exige beaucoup d’ eau .
Mais l’ ortie exige une terre non tassée ,ombragée , et riche en matière organique .Donc tout le contraire d’une zone agricole moderne.
Reference:
https://www.toutvert.fr/fil-dortie-avantages/
L’ortie , le coton du grand Est revue We-demain ( revue qui explore les techniques qui feront notre future ).
Dominique.
-
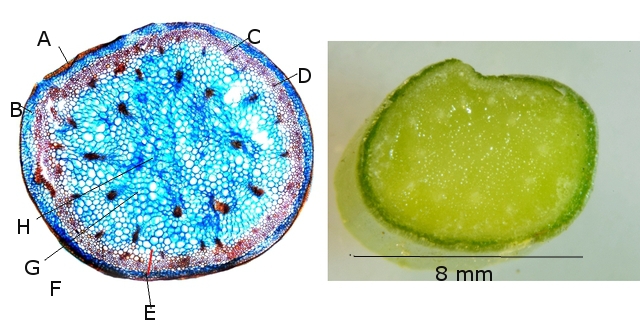
A –Epiderme.
B-- Parenchyme cortical.
C—Massif de Phloème.
D –Fibres lignifiées .
E –Parenchyme sclerifié ( sclérenchyme ).
F – Xyléme – faisceau cribro-vasculaire.
G –Parenchyme médullaire.
H --on constate qu il n y a pas de lacune centrale.
Le bulbe
La mise en réserve des nutriments nécessaires à la survie de la plante se fait dans les rhizomes et les bulbes -

Coupe d’un bulbe :

La coupe d’un bulbe permet la mise en évidence de ses différentes parties - ( qui sont plus visibles sur la coupe de l’ échalote ,photo de droite).
Le plateau épais et rond contient les cellules du méristème et qui est l’équivalent d’une tige.
Coupe de la colonne centrale ( partie initiale de la tige dans le médaillon ) .Son organisation est celle de la tige que l’ on a rencontré au début
Sur cette tige s’insèrent des feuilles.
Sur les côtés du plateau se développent les racines.
Coupe faite à hauteur du plateau.
Panorama de 50 photos (logiciel ICE)
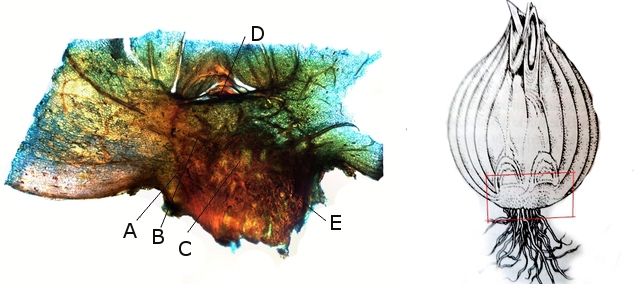
A –Initium foliaire (ébauche foliaire) qui apparaît progressivement sur le pourtour de l’anneau initial.
B –Centre générateur de feuilles.
C –Méristème médullaire (groupement de cellules totipotentes qui vont se différencier pour former les divers organes de la plante. Le méristème est responsable de la croissance en longueur des plantes. Le cambium est un autre groupement de cellules totipotentes .C’’ est un tissu de croissance secondaire situé sous l'écorce des plantes. Il est responsable de la croissance en épaisseur des plantes, produisant du bois et du phloème.
D –partie initiale de la tige.
E –naissance des racines qui dans le cas des liliacés se developpent sur la périphérie de l’ anneau initial.
Photo de gauche ( A ) départ du développement des racines ( primordium racinaire ).
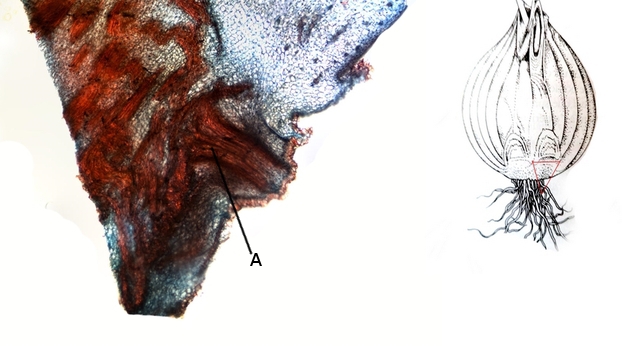
Ce primordium racinaire va à l’automne être à l’ origine de la formation des bulbilles - qui est la forme asexuée de la multiplication
C’est cette forme asexuée de multiplication qui est préférée par les horticulteurs puisqu’ il s’agit de clones gardant les caractéristiques de la plante parent .

Classification
Plantea
Régne Tracheobionta
Division Magnoliophyta
Classe Liliopsidar
Sous-classe Liliidae
Ordre Liliales
Famille Liliaceae
Genre Hyacinthus
Référence :
Manipulation de botanique de L Genévés Editeur Dunod
Cours de botanique générale de D Bach - M Mascré – G Deysson
Dominique



Lignification Topinambour Tige évolution au cours de l’année
dans Dominique
Posté(e)
Lignification Topinambour Tige évolution au cours de l’année
Le topinambour ( Helianthus tuberosus) est une plante de la famille des asteraceae bien connue Ces fleurs sont décoratives et ses tubercules sont comestibles et toujours présents sur les étals des marchés
Sa croissance est rapide au printemps la tige est très souple -Cette tige depasse les 2 métres en automne et devient particuliement rigide -
Comment expliquer cette rigidification ?
Coupe faite le 12 Mai - (la tige atteint les 15 cm de haut )
A--Epiderme
B –Parenchyme cortical
C –Cylindre central
D –Phloème
E --Cambium
F –Xylème
G –Rayons médullaires
H –Parenchyme medullaire
Coupe faite le 16 juin
Zone corticale
A—Epiderme sur lequel sont apparus des trichomes (A’)
B –Parenchyme cortical : son épaisseur a déjà beaucoup diminué.
C –Cylindre central. Il se situe de plus en plus vers la périphérie au dépend de la zone corticale
Axe vasculaire
D –Phloème
E –Cambium qui assure la croissance en centrifuge du Phloème et en centripète du Xylème
Zone médullaire
F –Rayons médullaires
G –Parenchyme médullaire
Coupe faite le 6 septembre
A--Epiderme
B –Parenchyme cortical :
Axe vasculaire
C –Phloème qui se lignifie
D –Cambium
E --Xylème
F –On note la disparition progressive des rayons médullaires cellulosiques qui sont remplacés par l’apparition de faisceaux collatéraux lignifiés ce qui augmente la rigidité de la tige .On noter aussi le début de lignification des rayons medullaires qui restent ( couleur rosé )
F –Xylème
Zone médullaire
G –Parenchyme médullaire qui tend à disparaitre dans son centre
Coupe faite le 5 Octobre (la tige atteint 1,90m)
Zone corticale
A—Epiderme Il s’est épaissi.
B –Parenchyme cortical : Il a quasiment disparu.
C –Cylindre central. Touche désormais l’écorce.
Axe vasculaire
D –Phloème .Il est totalement lignifié.
E –Cambium Il s’est aussi lignifié (le diamètre de la tige est désormais fixé et cette lignification augmente la résistance de cette tige.)
Zone médullaire
F –On note la disparition des rayons médullaires cellulosiques qui sont remplacés par des faisceaux collatéraux lignifiés ).
Conclusion
La rigidité de la tige est liée au développement progressif de la lignification Ce processus de lignification débute très tôt dans l’année avec les vaisseaux de Xylème et s’ étend au Phloème puis aux cambiums .
Par ailleurs en même temps que le parenchyme cortical se raréfiait sont apparus au cours de développement des faisceaux collatéraux qui se sont aussi lignifiés ( ces faisceaux collatéraux se sont développés au dépend des rayons médullaires cellulosiques qui sont de moindre résistance ).
La lignification est donc un processus évolutif tout au long du cycle de développement de la tige du topinambour.
Definition de Wikipedia
La lignine (du latin lignum qui signifie bois) est une biomolécule, plus précisément une famille de macromolécules polyphénoliques, qui est un des principaux composants du bois avec la cellulose et les hémicelluloses. Ce biopolymère est présent principalement dans les plantes vasculaires et dans quelques algues rouges coralligènes, ce qui suggère une convergence évolutive de la biosynthèse des lignines entre ces algues et les trachéophytes. Ses principales fonctions sont de conférer de la rigidité et de la résistance mécanique aux parois cellulaires, ainsi que d'apporter une imperméabilité à l'eau et une résistance à la décomposition.
****************************
Remarque technique sur cette observation :
Le protocole de coloration a toujours été le même mais les résultats ne sont pas stables Pour les 4 préparations on a procédé de la manière suivante et pourtant les colorations obtenues sont vraiment différentes.
Coupe autour de 120 µm
Bain dans un mélange alcool/ acide acétique / formol 10 minutes ‘(AFA).
Rinçage puis Eau de Javel du commerce durée jusqu’ au blanchiment des échantillons.
Rinçage coloration avec un mélange (solution de safranine 3 gtt + eau 6 gtt + Solution d’Acridine 2 gtt ) 3 minutes
Rinçage puis bain dans une solution d’Alun de Potasse 7 % 3 minutes
Rinçage coloration avec le mélange (Astra Bleu 2 gtt + 6 gtt d’eau + Bleu Alcian 2 gtt ) 2 minutes
Déshydratation avec un mélange Alcool 95 + Fotonal 1% (pour éviter les bulles d’air)
Xylène quelques gouttes sur la lame (pour éviter les bulles d’air)
Montage Euparal
Explication possible un manque de rigueur dans les temps de manipulation des échantillons lors des différentes étapes ( et peut-être 1 goutte de plus ou de moins par mélange - mais la manipulation de la pipette Pasteur ‘n’est pas aussi évidente qu’ il y paraît ) .
Dominique.