Dominique.
-
Compteur de contenus
1 481 -
Inscription
-
Dernière visite
-
Jours gagnés
1
Tout ce qui a été posté par Dominique.
-

Plante carnivore Sarracenia leucophylla
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
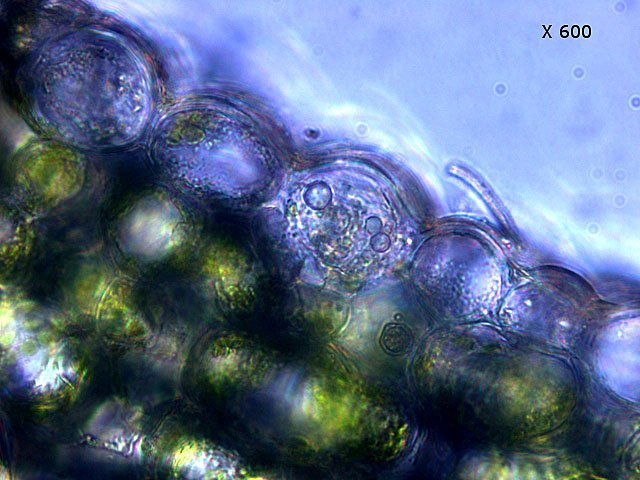
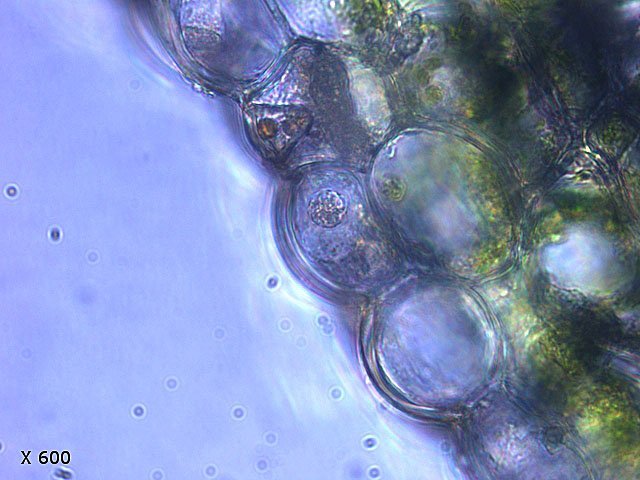
Il faut noter que la plante n’ arrive pas à digérer la chitine de l’ exosquelette des insectes - qui vont doucement encombrer l’ intérieur de l’ urne ( dans le cas de cette observation les urnes sont vides - ce sont des plantes de jardinerie vendues à un âge précoce et vivant dans un milieu quasi aseptisé Cet aspect spectaculaire des plantes carnivores ne doit pas cacher le fait que la principale source de protéines est fournie par la synthèse chlorophyllienne :Les cellules de cette feuille contiennent toutes des chloroplastes. Grossissement X600 Dic: Discussion *************** Dans plusieurs articles il est dit que cette feuille a été obtenue par un enroulement du limbe , suivit de la soudure des deux bords -Cette explication permettrait de rendre compte de la situation de la face supérieure d’ une feuille plate qui ici représente la face interne du tube Les deux coupes suivantes s’opposent à cette affirmation Ces deux coupent sont réalisées un peu au- dessus du niveau de la photo numéro 3 de cet article. Elle met en évidence que le tube se forme dès la base . --Dans un premier temps il y a disparition du parenchyme lacuneux de la nervure principale . --Dans cet espace la différentiation cellulaire va aboutir à fabriquer un épithélium et un parenchyme cortical identiques à ceux de la face supérieure d’une feuille (Pour la synthèse et l’ organisation de cet épithélium le code génétique n' a donc pas été modifié ). --Le tube ainsi organisé va progressivement s' allonger pour atteindre sa taille définitive . Il n’y a donc pas d’enroulement ni de creusement Quelques informations sur les Sarracénies ( Ref 1 ) Les Sarracénies ou Sarracènes (le genre Sarracenia) sont un genre de plantes à fleurs de la famille des Sarracéniacées. Ce sont des plantes carnivores originaires du sud-est des États-Unis et du Canada. Ce sont des « carnivores à pièges passifs » (sans mouvement mécanique). Ces pièges sont des feuilles modifiées en forme d'urne et protégées par une coiffe, sauf chez Sarracenia psittacina chez qui elle a disparu par soudure de ses bords. Le nom de la plante fait référence à Michel Sarrazin, chirurgien, médecin et naturaliste en Nouvelle-France1. Description et caractères À l'extrémité apicale, des glandes nectarifères sécrètent un suc qui attire les insectes en plus de la synthése d’une pheromone la Sarrcenine Il a été montré que les frelons asiatiques sont particulièrement attirés par cette molécule dont on essaie de faire la synthèse. Pour les sarracénies à urnes verticales, la coiffe ne sert pas à refermer le piège mais à éviter que l'eau n'y rentre. Chez Sarracenia minor elle empêche aussi les insectes de ressortir ce qui fait que les poils obliques sont absents. Chez Sarracenia purpurea, les urnes sont couchées et la coiffe facilite au contraire la récupération de l'eau de pluie. L'eau est nécessaire pour piéger par noyade les insectes et gastéropodes mais facilite également la digestion. Les sarracénies produisent des fleurs qui apparaissent en général à partir de leur quatrième année, au printemps pour les précoces Sarracenia flava et Sarracenia oreophila, jusqu'au milieu de l'été pour Sarracenia minor et Sarracenia leucophylla. Classification Règne Plantae Division Magnoliophyta Classe Magnoliopsida Ordre Nepenthales Famille Sarraceniaceae Genre Sarracenia L , 1753 Références https://fr.wikipedia.org/wiki/Sarracenia_leucophylla https://forum.MikrOscOpia.com/topic/14718-sarracenia-purpurea-plantes-carnivores-1/#comment-54270 article de Pierre https://www.futura-sciences.com/planete/dossiers/botanique-plantes-carnivores-478/page/4/ https://positivr.fr/sarracenia-plante-carnivore-mange-frelons-asiatiques/ https://www.kloranebotanical.foundation/la-botanique/dossiers/plantes-carnivores/comment-mangent-les-plantes-carnivores https://pubmed.ncbi.nlm.nih.gov/35604105/ digestion https://www.carnivorousplants.org/cp/carnivory/digestion https://academic.oup.com/plphys/article/190/1/44/6590657 Dominique.
-
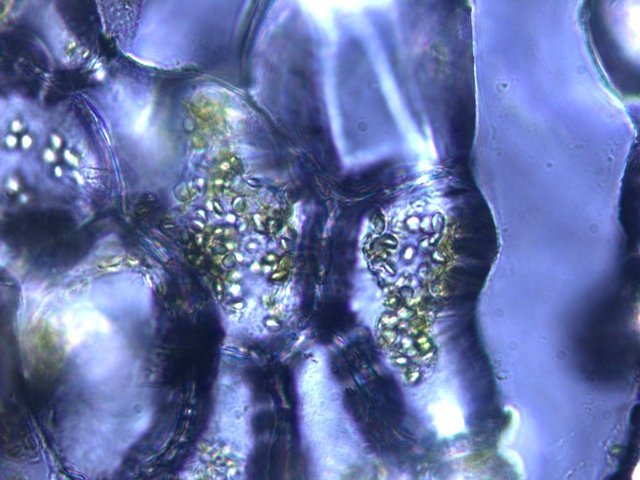
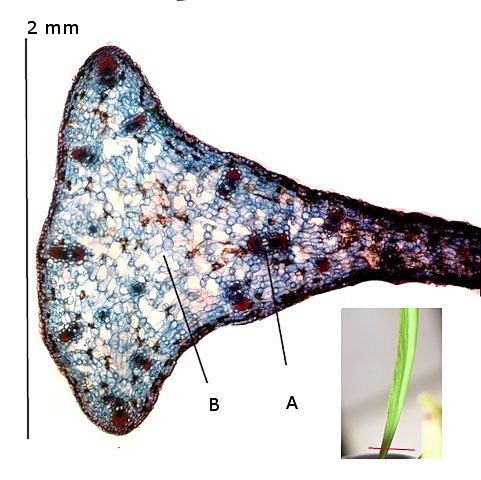
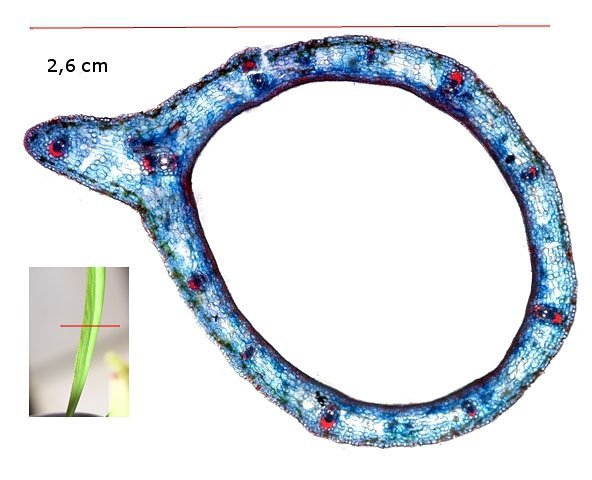
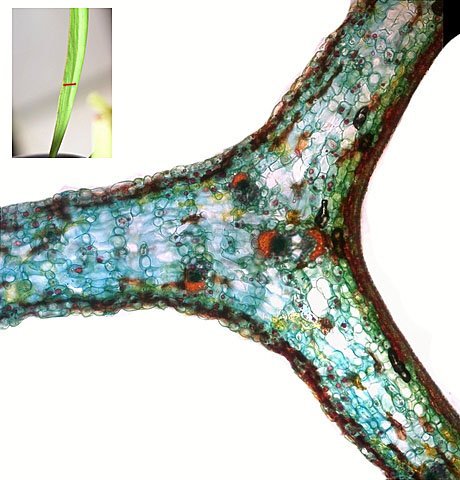
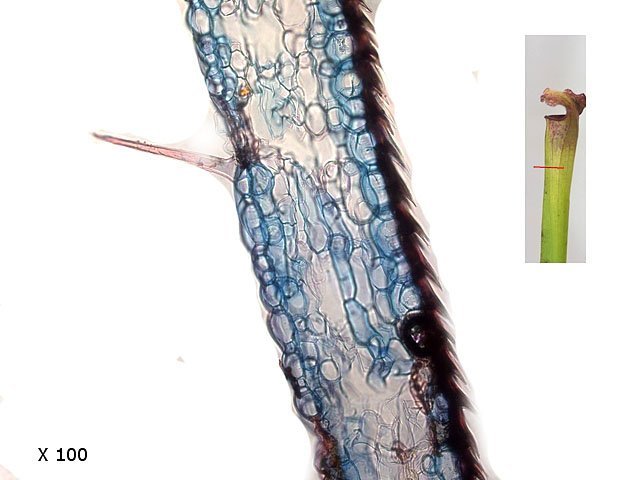
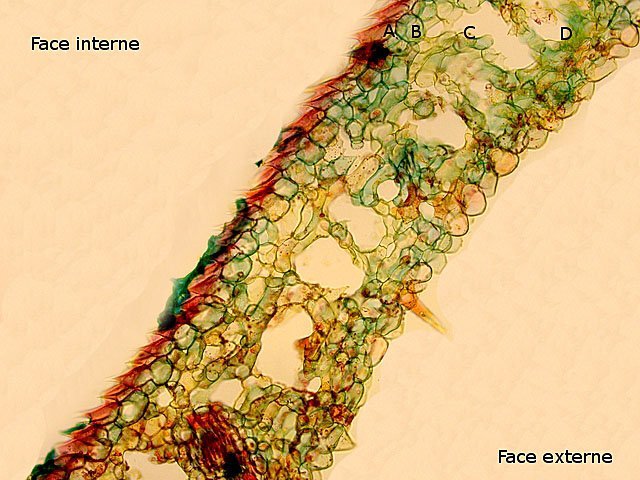
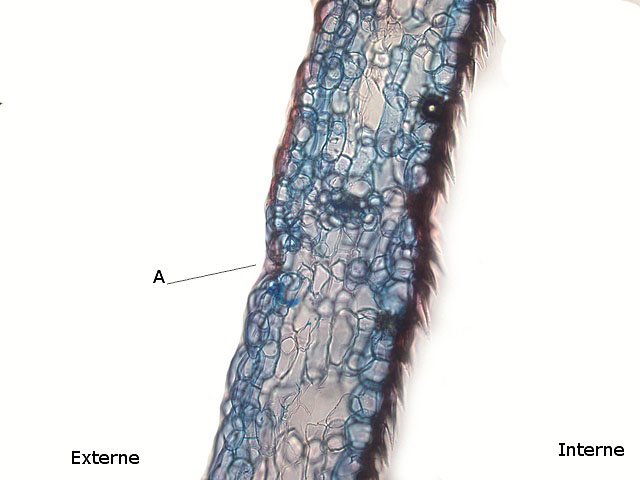
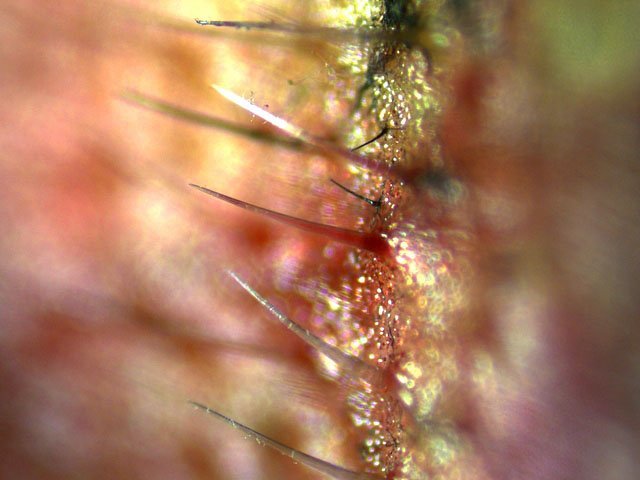
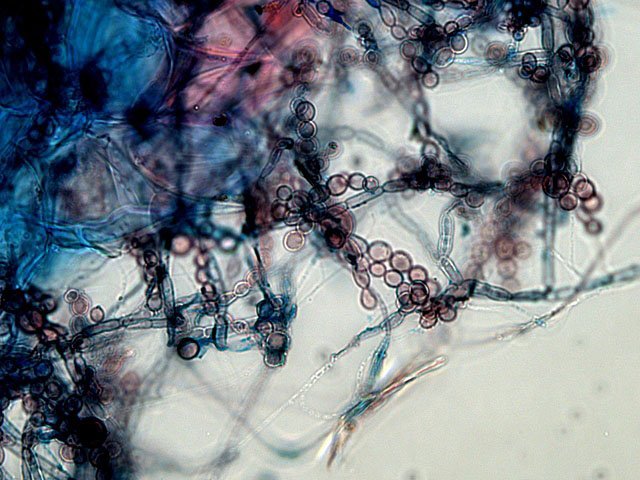
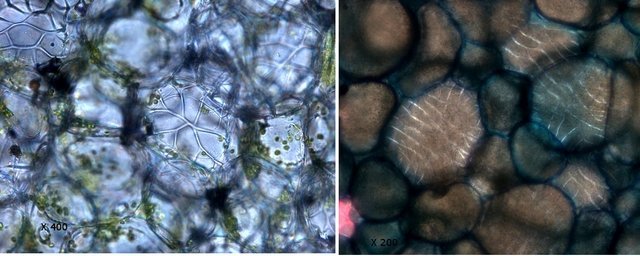
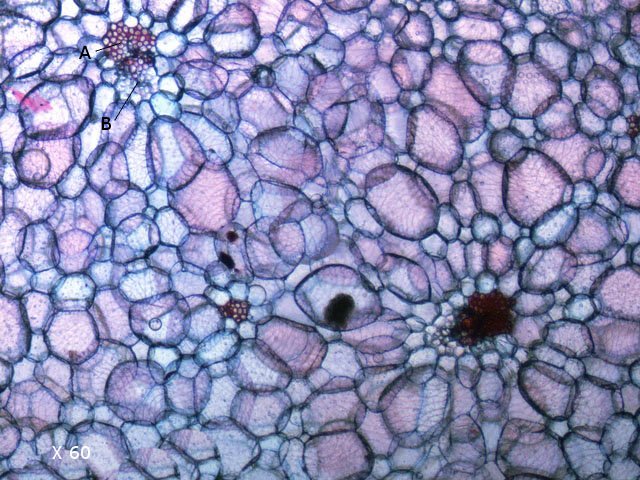
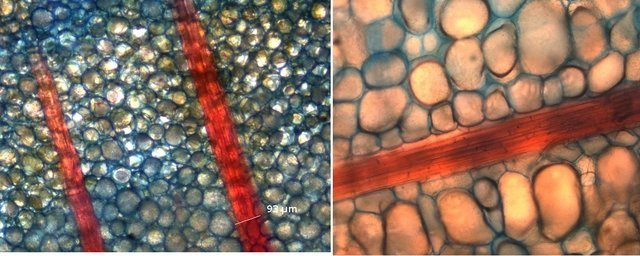
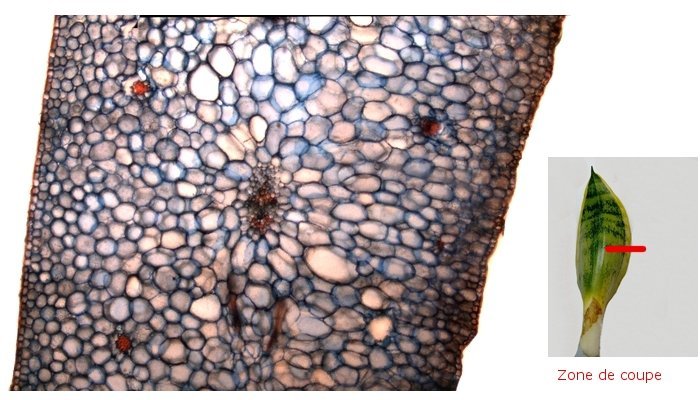
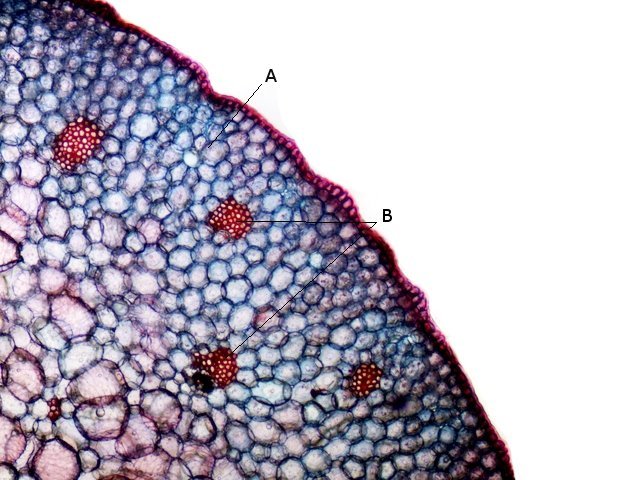
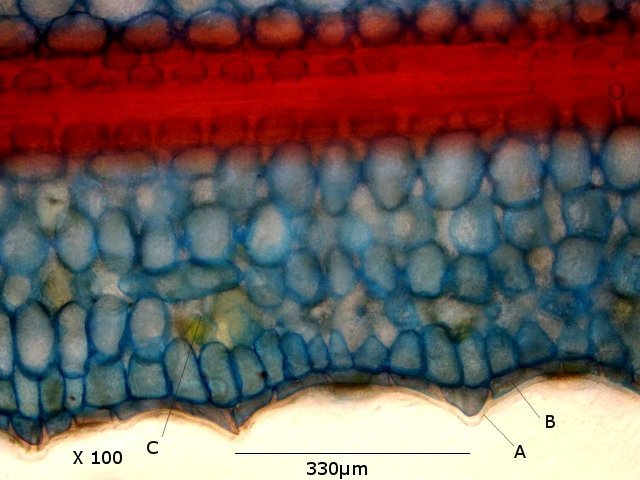
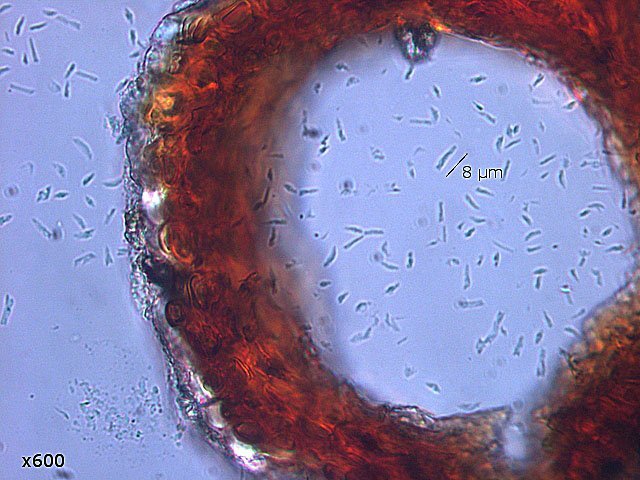
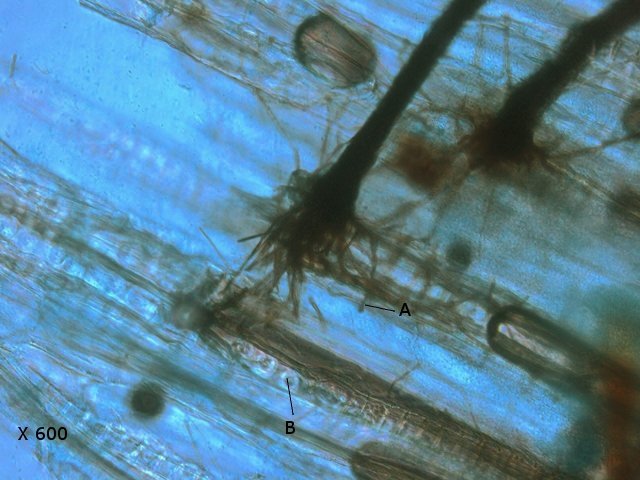
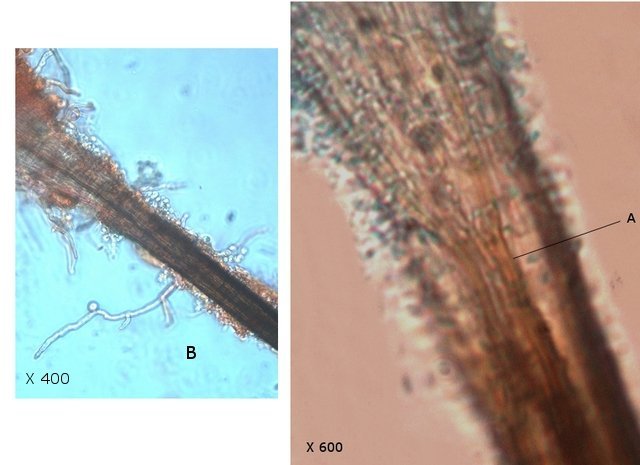
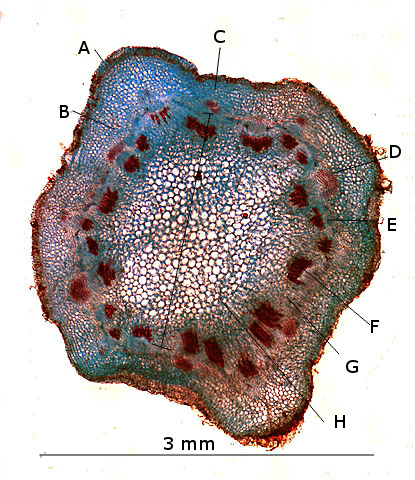
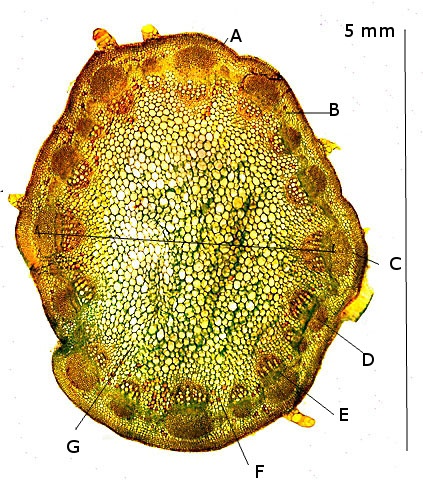
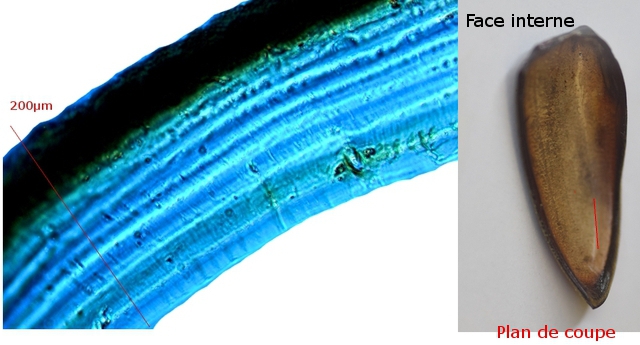
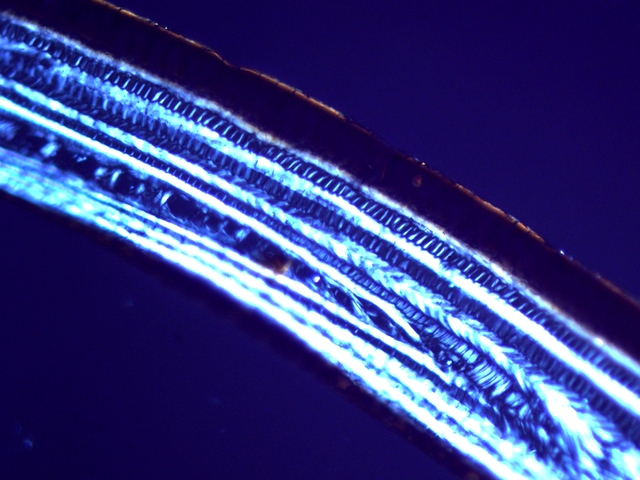
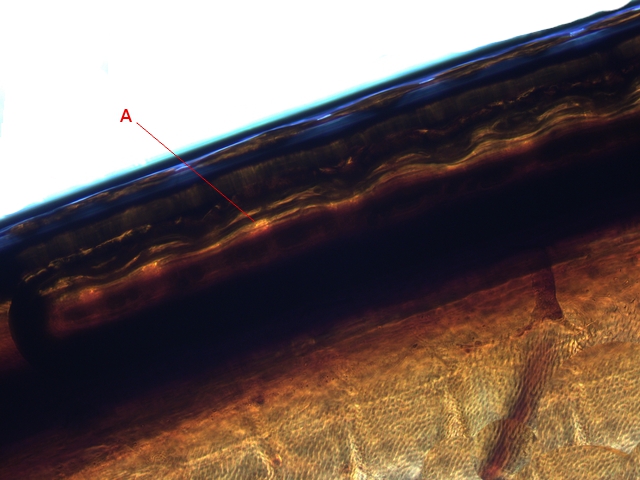
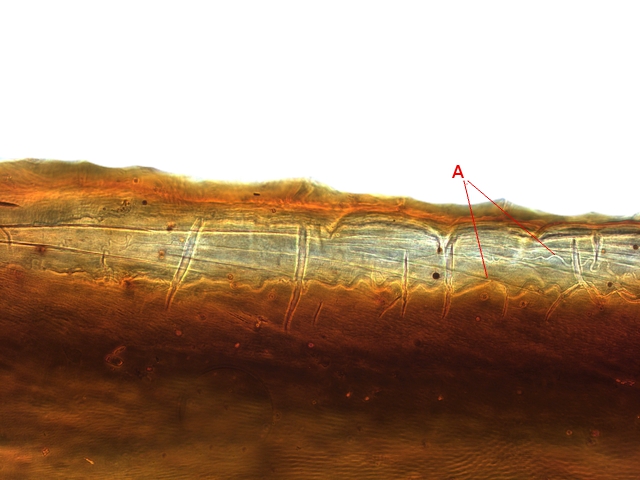
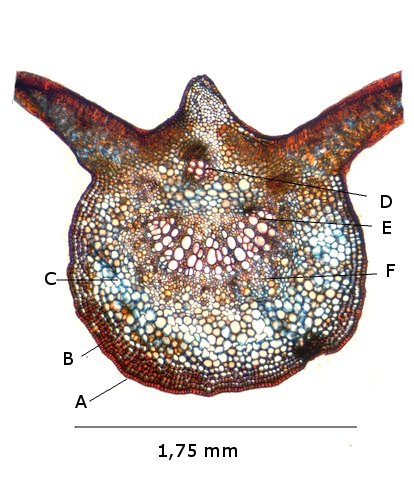
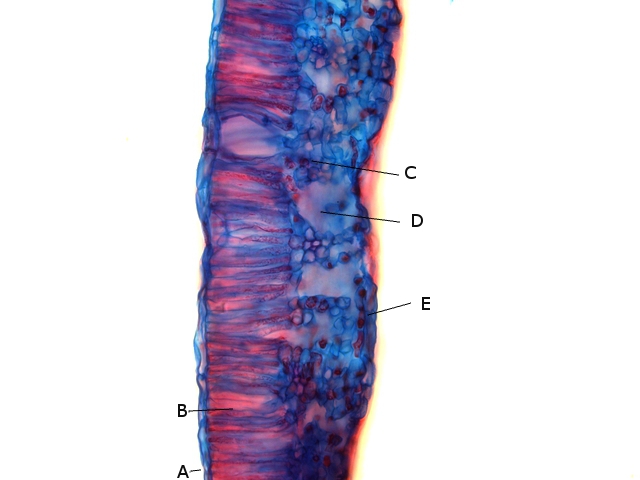
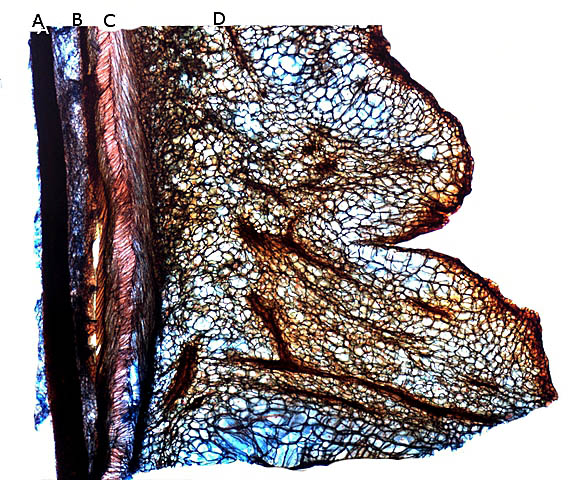
Plante carnivore Sarracenia leucophylla Poursuite de l’étude des feuilles https://forum.MikrOscOpia.com/topic/16067-feuille-histologie-consistance/#comment-62043 https://forum.MikrOscOpia.com/topic/19892-sansevieria-cylindrica-%C3%A9tude-histologique/#comment-82518 Il a déjà été présenté des feuilles plates ( les plus fréquentes ) les feuilles cylindriques ( Sensevieria cylindrica ) ;cette fois ci on se penche sur les feuilles tubulaires Les plantes carnivores ont transformé leurs feuilles en piège La Dionée est une plante carnivore à piège actif qui referme rapidement ses « mâchoires » autour de sa proie ; les Nepenthes sont dotées d’un piège passif formé d’urnes remplies de liquide dans lesquelles les insectes se noient ; le Drosera est pourvu d’un piège semi-actif sous forme de feuilles collantes desquelles les insectes ne peuvent plus se dépêtrer. Chez Sarracenia leucophylla le piège est un tube où vont tomber les insectes comme chez les Nepenthes Cette plante peut se trouver facilement en jardinerie ( mais dans ce contexte la plante est de petite taille ). La morphologie en tube n’existe pas dès le début de la formation des feuilles -mais la forme définitive de la feuille va s’installer rapidement. . Pour comprendre l’organisation de cette feuille il est réalisé des coupes étagées de bas vers le haut. Partie basse Dans cette partie le tube n’est pas encore constitué: A – Axe vasculaire B – Parenchyme lacuneux Cet aspect de la base de la feuille sera discuté plus tard Coupe partie moyenne: Coupe au niveau du tube. Cette coupe montre qu’ à l’origine de la courbure il existe un axe vasculaire ( A ) comme habituellement présent dans les nervures des feuilles .La zone de division est donc la nervure principale de la feuille - par contre dans cette observation il n’ existe pas de nervures secondaires ni de nervioles . Organisation d’une feuille : L’ organisation de la répartition cellulaire de cette feuille est en fait classique: A Cuticule B Parenchyme cortical Il existe une seule couche de cellules épithéliales avec un faible développement du parenchyme palissadique C on constate un fort développement du parenchyme lacuneux qui représente une grande partie du parenchyme D il existe très peu d’éléments ligneux secondaires En fait on se trouve dans une architecture très proche de la feuille de nénuphar Par ailleurs on constate si on se réfère à la structure la plus fréquemment rencontrée chez le feuilles que la face supérieure de la feuille forme la face interne du tube et que la face inférieure forme la partie la plus externe de ce tube. Il faut en conclure que la feuille s’est développée comme si elle s’était enroulée sur elle-même avec soudure des bords ( la face supérieure de la feuille est donc devenue interne ) - Cette interprétation est fausse on le verra plus tard. Sur la face externe il existe des stomates en connexion avec les zones aériennes de la feuille - La fonction piége : Le piège est à la fois chimique et mécanique : Le couvercle de la tige est porteuse d’une grande quantité de poils : L’ entrée de l’ urne est le lieu de synthèse d’une molécule attractive la Sarracenine Coupe du bord de la feuille qui révèle que ce bord se recourbe sur lui même: Une fois l’insecte engagé dans le tube le retour à reculons est entravé par un ensemble de poils raides dont l’ orientation s’oppose à tout retour ( comme les dents de la couleuvre ). Le tube des Sarracenia crée un biotope ; La digestion est le résultat de plusieurs actions. A Les bactéries Une fois l’insecte bloqué dans le tube la digestion se fait en grande partie par les bactéries. B Les moisissures L’ examen des cellules de la paroi interne de l’urne met en évidence la présence de différentes moisissures qui sont situées dans cette observation surtout dans la partie inférieure du tube. C La digestion est aussi assurée par la synthèse d’enzymes .l’ examen des cellules de la plante met en évidence la présence de nombreux Leucoplastes .
-
Araignée de mer étude de son système optique Maja brachydactyla
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
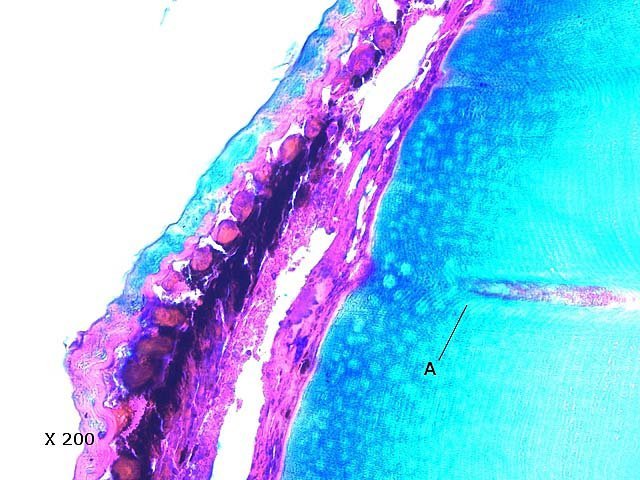
Bonsoir Si on reconsidère les images en faisant abstraction de la structure des ommatidies des insectes avec un cristallin pour chaque ommatidies les formations rondes ( B ) de la photo 8 et celle de la formation de la photo 9 peuvent être considérées comme une coupe perpendiculaire des tubes de l’ œil Si on regarde attentivement la photo 9 chaque rond est limité par une paroi bien visible. Il est possible de lire cette image: a cellules pigmentaires périphériques b cellule rétinienne c rhabdome (la zone transparente centrale ou passent les rayons lumineux. Pour le cristallin sur la photo 8 on constate que sa surface interne est bosselée - et que chaque convexité correspond au départ de quelque chose qui pourrait être un tube. ( par contre le cristallin n' est pas individuel à chaque unité de vision comme pour les insectes ) En fait la difficulté d’interprétation vient en grande partie des plans de coupe de l’échantillon. Il sera donc nécessaire de refaire ces coupes pour être certain de la bonne interprétation; mais de toute façon on se trouve bien devant un œil . composé Amicalement Dominique
-
Araignée de mer étude de son système optique Maja brachydactyla
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
Bonsoir Tryphon et Solito de Solis Merci pour toutes ses informations ( et le livre de ED Yong ) .Je vais essayer de contacter une des stations de biologie marine . Il y a une station Ifremer à quelques kilomètres à Port en Bessin . Je vous tiendrai au courant Amicalement Dominique -
Araignée de mer étude de son système optique Maja brachydactyla
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
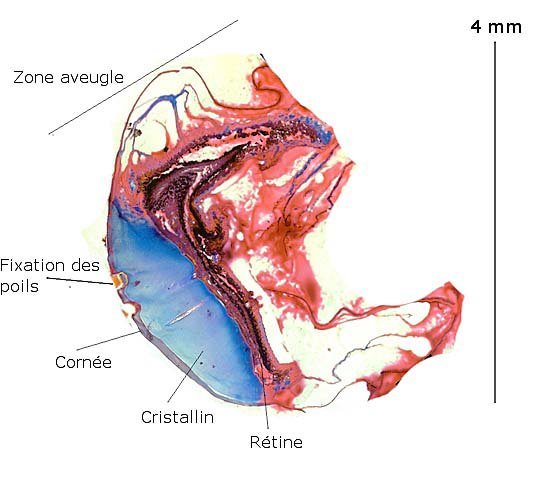
Bonjour Jean -Marie L’œil dans sa partie supérieure est protégé par la carapace ( que j' ai dissous pour faire les coupes ) cette zone est la zone aveugle. Pourtant en dessous de cette zone couverte par la carapace il existe toujours une membrane rétinienne qui s' est pliée ( et qui ici ne sert semble t il à rien )- Il y a probablement eu un processus évolutif la carapace protégeant l' œil de façon plus efficace. La forme alvéolée représente la totalité de la cornée . La encore pourquoi cette forme alvéolée alors qu" il n' y a pas d' ommatidies derrière -Comme si l' évolution avait conduit à la formation d'une rétine unique tout en gardant la structure alvéolée de la cornée primitive . ( cette interprétation est toute personnelle ). Amicalement Dominique -
Araignée de mer étude de son système optique Maja brachydactyla
Dominique. a posté un sujet dans Dominique
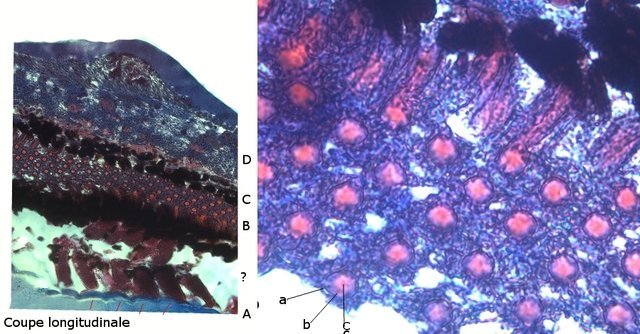
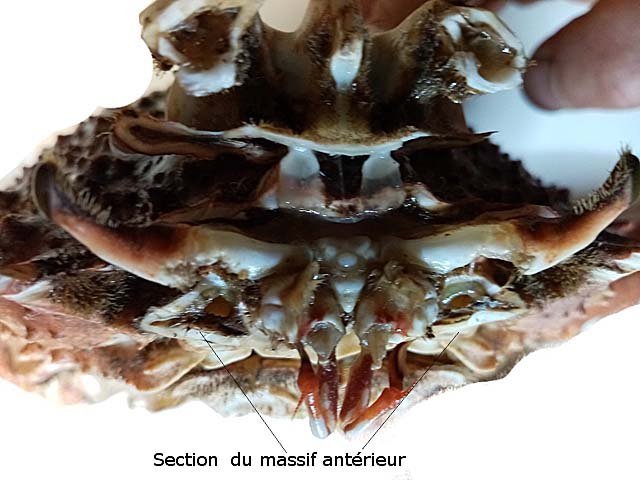
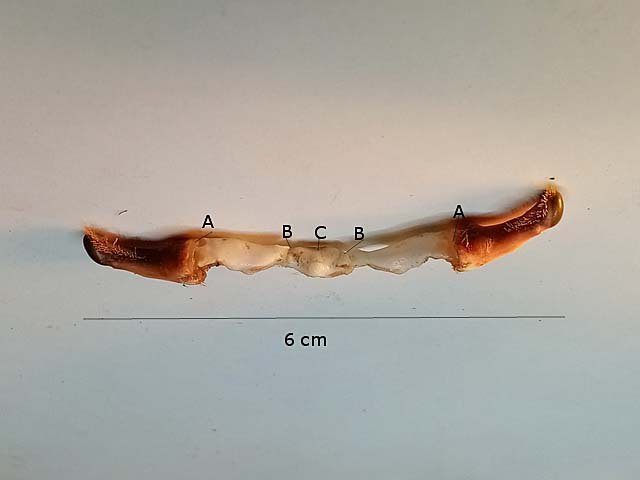
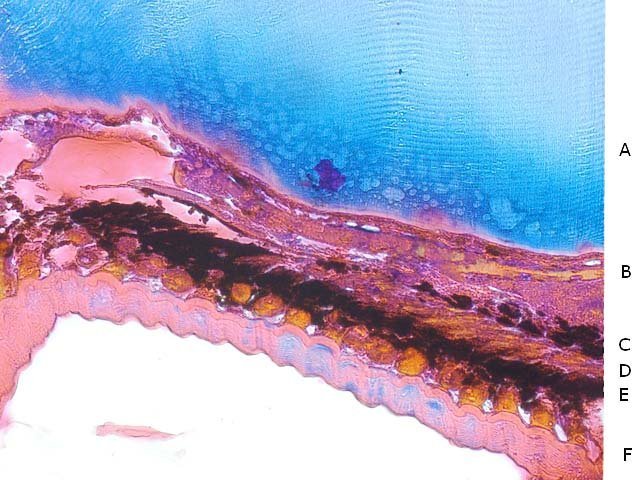
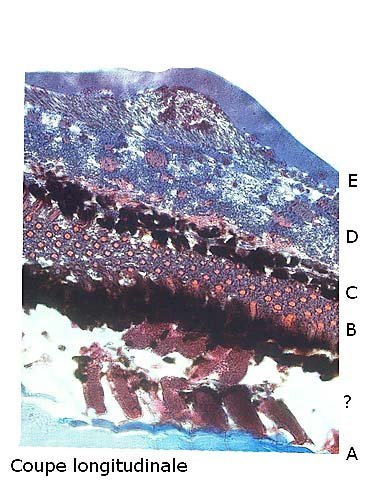
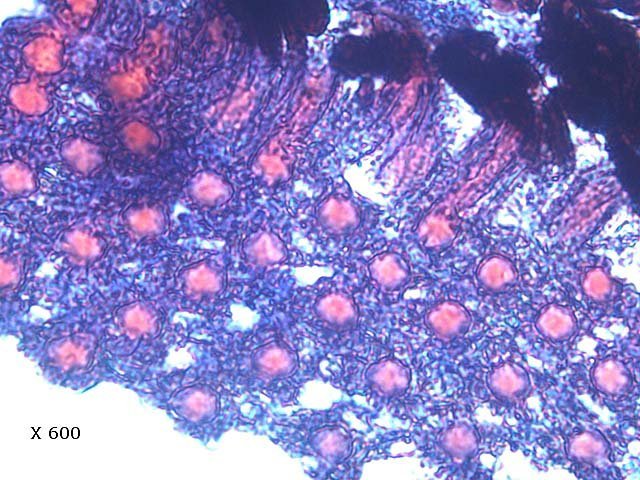
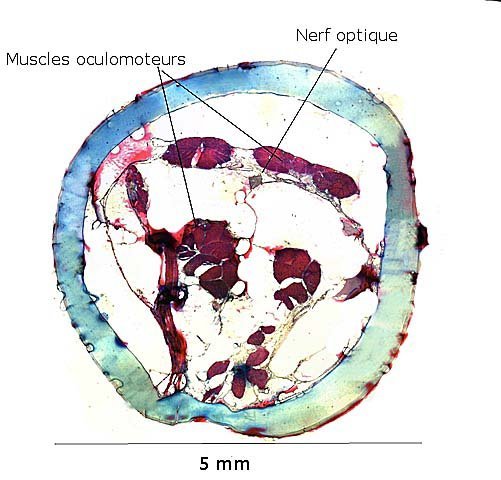
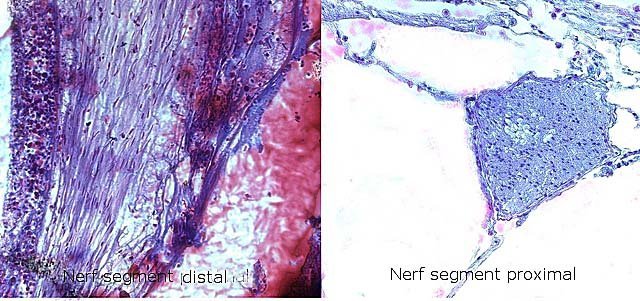
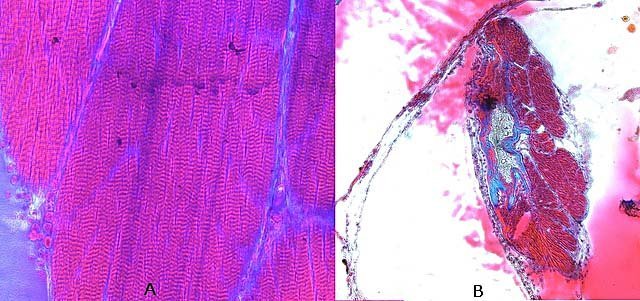
Araignée de mer : étude de son système optique. Maja brachydactyla Sur le marché l’ étale du poissonnier présente surtout 2 types de crabes - des tourteaux et des araignées de mer -Ce qui est frappant chez ces animaux ce sont leurs yeux en grande partie cachés par leur carapace qui de ce fait semblent saillir de part et d’autre . La partie antérieure de la carapace est découpée .ce qui permet de mettre en évidence la totalité du système optique dans son logement une fois la zone antérieure de la carapace réclinée. Les yeux sont donc situés ce chaque côté à l’ extrémité de deux segments ( les pédoncules ) Le segment proximal (B ) s’articule par son extrémité interne sur un axe central ( C ) qui est immobile Le segment distal ( A ) s’articule pas son extrémité interne avec l’ extrémité distale du segment (B ) .Son extrémité externe porte le système optique. Il est constaté que l’œil de l’ araignée de mer est orienté vers le sol et que de ce fait sa vision vers le haut est limitée même avec les déplacements qu’ autorise cette double articulation Si on réalise une photo épiscopique ( X 40 ) de cet œil on constate que la surface de la cornée est en aspect alvéolée comme le sont les yeux des arthropodes qui possèdent des yeux composés A noter aussi la présence de poils: On peut donc lire dans de nombreuses publications Les yeux composés existent uniquement chez certains insectes et crustacés. Ils sont constitués d'un grand nombre d'unités visuelles : les ommatidies. » Ce que mes coupes ne mettent pas en évidence Voilà ce qui est constaté L’examen de l’ œil des crustacés de type crabe ne peut se faire que si on décalcifie les échantillons avant de réaliser des coupes Les coupes ont été colorées au trichrome de Gilles Hematoxyline de Gilles – Eosine - bleu d’ Aniline Vue générale On constate donc -- cet œil n’est pas un œil composé comme celui des insectes. -- le cristallin est extrêmement épais -- les encoches dans la zone superficielle du cristallin sont en lien avec l’insertion des poils vus en épiscopie -- la partie profonde est constitué d'une couche pigmentée. Examen d’une division en profondeur En A ces divisions affectent la courbure des fibres du cristallin Elles ont une disposition irrégulière et ne définissent aucun cône ( comme pour les ommatidies des insectes ) par contre elles doivent être à l’origine d’ une diffraction de la lumière par la modification des courbures des fibres du cristallin . La rétine La rétine est constituée de plusieurs couches tissulaires Coupe axiale A – Cristallin B – Tissu gliale C - Ce qui frappe est l'organisation régulier d' éléments juxtaposés .Cette image évoque un œil composé D – Cellules photo réceptrices ou plus probablement couche nucléaire des cellules réceptrices E – Epithélium pigmentaire F Choroïde et Sclére Coupe longitudinale A –Cristallin B Cellules photo-réceptrices L’organisation sur cette photo est faite d’unités arrondies régulièrement espacées Cet aspect est très évocateur des ommatidies Ces ommatidies sont des unités cylindriques .C’est sur la paroi de ces tubes que se développent les cellules photoréceptrices : les cellules de la rétine . Ce type d’ organisation est typique des yeux des insectes C –Cellules pigmentaires D – Choroide E – Sclére ( Sclérotique ) Examen de la zone réceptrice au X 600 C –Cellules pigmentaires D – Choroide E – Sclére ( Sclérotique ) Examen de la zone réceptrice au X 600: Coupe des ommatidies En fait chaque point rouge correspond a un œil .Chaque tube est tapissé avec les cellules de la rétine La lumière concentré par le cristallin va pénétrer dans ces tubes Les photons de la lumière vont activé les cellules photo réceptrice qui vont transmettre l’ information au cerveau ,lieu où va se créer l’ image Par comparaison Œil du poisson https://forum.MikrOscOpia.com/topic/17094-%C5%93il-poisson/#comment-67863O Œil de la crevette https://forum.MikrOscOpia.com/topic/19097-crevettes-2-%C3%A9l%C3%A9ments-d%E2%80%99anatomie/#comment-79200 Œil du calamar https://forum.MikrOscOpia.com/topic/19637-calamar-l%E2%80%99%C5%93il/#comment-81523 La différence d’ organisation est frappante en comparaison avec l’ œil de l’ araignée de mer Le caractère moins sophistiqué chez celle-ci est évident Qu’ en est il de la qualité fonctionnelle ? De toute façon cette qualité est suffisante pour la survie de l’ animal qui vit dans les océans depuis le Cambrien soit il a y 500 millions d’ années Organisation des segments mobiles de l’œil ( Les Pédoncules ) Les deux segments mobiles du système optique de l’ araignée de mer sont de même structure – Coupe du segment proximal . ( X 40 ) Les muscles oculomoteurs vont permettent l’orientation de l’ œil – On compte 6 massifs musculaires Ce fait est à rapprocher de l’ œil humain qui possède six muscles oculomoteurs assurant les mouvements de chaque globe B Le nerf optique Dans le segment distal ( donc qui se termine par l’ œil ) le nerf optique n’ est pas organisé et forme un plexus . Ce qu’il n’ est pas le cas dans le segment proximal . A ce niveau l’organisation en nerf est réalisée avant de pénétrer la carapace et de rejoindre le Protocerebrum A la sortie de la sclérotique Avant sa connexion avec le Protocerebrum Les muscles oculomoteurs sont des muscles striés. A – Les muscles sont de type striés B – Il est possible d’ individualiser un système vasculaire qui coure le long des muscles Règne Animalia Embranchement Arthropoda Sous-embr. Crustacea Classe Malacostraca Sous-classe Eumalacostraca Super-ordre Eucarida Ordre Decapoda Sous-ordre Pleocyemata Infra-ordre Brachyura Super-famille Majoidea Conclusion Reste le probléme du fait que je ne constate pas d’ œil composé comme celui des insectes alors que beaucoup d’ autres publications ne font que relater le fait L’œil composé est bien particulier comme on s’en rend compte dans l article suivant portant sur le cerveau du frelon asiatique https://forum.MikrOscOpia.com/topic/16406-frelon-asiatique-cerveau-histologie/#comment-64192 Reference https://www.sciencedirect.com/ science/article/abs/pii/S1467803906000521 Dominique. Et bonne année à tous
-
Sansevieria cylindrica étude histologique
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
Bonjour Il est vrai que les microtubules ne sont pas visibles au microscope optique - Cette affirmation a été tirée de l' article de référence portant sur l' histologie du Sansevieria. Par contre une accumulation de microtubules peut être visible . Amicalement Dominique -
FORMES LARVAIRES DU PLANCTON MARIN
Dominique. a répondu à un(e) sujet de Jean Marie Cavanihac dans Jean Marie CAVANIHAC
Bonsoir Jean -Marie Pour te remercier de la publication depuis 3 semaines de toutes ces belles planches qui sont le résultat d'un magnifique travail. Amicalement Dominique. -
Sansevieria cylindrica étude histologique
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
Sur cette coupe longitudinale on constate que les cellules sont organisées en colonnes verticales sur l’ensemble du parenchyme cortical et médullaire. Et en particulier de grosses cellules qui semblent faire des pylones . Particularité des cellules parenchymateuses du Sansevieria Les cellules se « développent au milieu d’un maillage fibreux ( microtubules ) Ces fibres sont bien visibles en lumière polarisée Aspect du parenchyme médullaire ( zone centrale ). A Xyléme B Phloéme Les vaisseaux sont nombreux selon une trajectoire ascendante Coupe longitudinale : Reste à répondre à la question de départ ( comment expliquer la position des feuilles du pot vendu en jardinerie ?) Si on regarde la description de Sansevieria sur un article de Wikipedia il est noté que cette plante peut atteindre 1 mètre Toutes les plantes vendues font 20 cm maximum Par ailleurs la disposition des feuilles est des plus curieuses : chaque feuille semble indépendante comme on l’avait noté au départ La solution est apportée par le dépotage de la plante : Ce qui est vendu est le résultat du bouturage de coupe de la partie haute de la feuille d'une plante déjà bien developpée .Dans la pot vendu il y a 6 extrémités de feuille. L’enracinement s’ est très bien fait avec le mélange de terre qui a permis à ces pousses de développer un racinaire. La multiplication peut se faire par division du rhizome ou par boutures. Les fragments doivent mesurer au moins 7 cm de long et être insérés dans un sable humide. Feuille ou tige ? Faisons une comparaison avec la feuille et la tige de la tulipe: On remarque une similitude d’organisation entre la tige de la tulipe et la feuille de Sansevieria cylindrica Comparons maintenant la structure d’une feuille de Sansevieria trifaciata qui a une forme plate: On constate que son organisation est la même que celle de la variété cylindrica Conclusion Quand on reprend le texte d’une étude scientifique (‘ Reference 1 ) il est parlé de feuilles Dans les traités de botanique il est dit que les feuilles présentent une diversité morphologique considérable ce qui est donc constaté ici puisque l’ aspect de cette feuille est quand même très proche de la structure histologique d’une tige Classification Règne Plantae Sous-règne Tracheobionta Division Magnoliophyta Classe Liliopsida Sous-classe Liliidae Ordre Liliales Famille Liliaceae Genre Sansevieria Espèce Sansevieria cylindrica (Wenceslas Bojer), 1837 Classification APG III (2009) Et pour conclure la fleur de la plante ( image extraite du web ) On remarque tout de suite que les feuilles de cette plante ne sont pas disposées comme les feuilles vendues dans la jardinerie. Dans le cas ci-dessus il ne s’agit pas de boutures mais bien de feuilles qui se sont développées sur un rhizome. Leur caractère cylindrique n’apparait que quelques centimètres au-dessus du pied Donc l’étiquette que portait ce pot de fleur était fausse ;-elle aurait dû être nommée Pot de 6 boutures de Sensevieria Cylindrica CQFD A noter que cette pante possède des propriétés pharmacologiques qui lui confèrent des effets antibactériens, anti-inflammatoires, analgésiques et antipyrétiques. Les études scientifiques ont montré que les extraits de Sansevieria cylindrica sont efficaces contre une variété de bactéries, notamment Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa et Streptococcus pneumoniae. Ils sont également capables de réduire l'inflammation, la douleur et la fièvre. En outre, la Sansevieria cylindrica a des propriétés antioxydantes qui peuvent aider à protéger les cellules contre les dommages causés par les radicaux libres. Références : https://www.phcogres.com/sites/default/files/PharmacognRes-14-4-412.pdf Dominique
-
Sansevieria cylindrica étude histologique Au cours d’une visite à la jardinerie de la ville j’ ai été surpris par la forme d’une des plantes - Elle ne présentait que des tiges dressées sans feuille apparente Son nom indiqué sur l’étiquette : Sansevieria cylindrica Le Sansevieria trifasciata est bien connu dans les maisons il fait partie des plantes d’ appartement souvent rencontrées en Normandie - Mais dans ce cas ses feuilles sont plates Dans le cas présent les feuilles sont cylindriques Une question se pose d’emblée pourquoi cette disposition de feuilles parallèles dès la sortie de la terre. ? Cette présentation porte la structure histologique de cette plante . Coupe de la feuille cylindrique : Les coupes sont très faciles à bien réaliser Coupe en partie haute (diaporama de 70 photos au X 80) Coloration bleu d’ aniline et safranine A Les Sclerenchyme lignifé est en périphérie - il n’existe pas de cylindre central bien dessiné B Présence fibres lignifiées dans le plan transversal C Axes vasculaires Xyléme et Phloem Contrairemenr aux autres feuilles il n’ existe pas de parenchyme spongieux qui d’ habitude forme des chambres ouvertes vers l’ extérieur pour permettre les échanges gazeux à travers les stomates et contrairement aux tiges il n’ y pas non plus de parenchyme médullaire Donc le mésophile ( partie moyenne de la feuille est homogène . Cependant première constation est que ce type de structure est plus proche de la structure d’une tige que celle d’une feuille L’épiderme est couvert de stomates: A cellules stomatiques (dites de garde ) B ostiole qui ouvre dans la chambre sous stomatique Sur ces deux photos il est noté le caractère très épais des cloisons intercellulaires: Les chloroplastes La photosynthèse est assurée par les Chloroplastes ( contenant la chlorophylle ) qui sont très nombreux dans les cellules les plus externes du parenchyme cortical Organisation de la zone corticale: A Cellules corticales - on constate qu’ il n’ existe pas de cellules palissadiques comme sur les autres feuilles et que toutes les cellules ont le même aspect - bien rond B Axe de sclerenchyme La cuticule La cuticule est la couche protectrice de la plante vis-à-vis des agressions extérieures. A cuticule B cellules épithéliales C cellules du parenchyme cortical La cuticule est formée de deux couches A cire qui forme la couche la plus externe B cutine et subérine C semble apparaître par places des lacunes aériennes. La rigidité de cette plante Cette rigidité est assurée par la présence de faisceaux fibreux correspondant à la une forte lignification du sclérenchyme Coupe longitudinale: La structure du parenchyme central est très homogène puisqu’ il ne présente des cellules arrondies de taille variable - Ce qui est frappant est leur organisation :
-
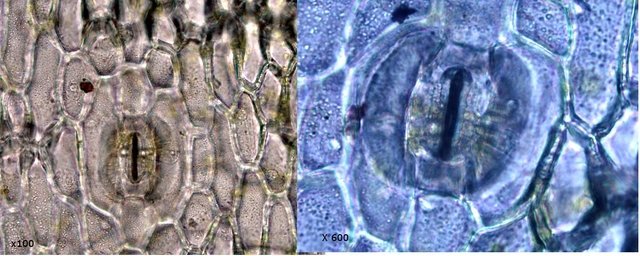
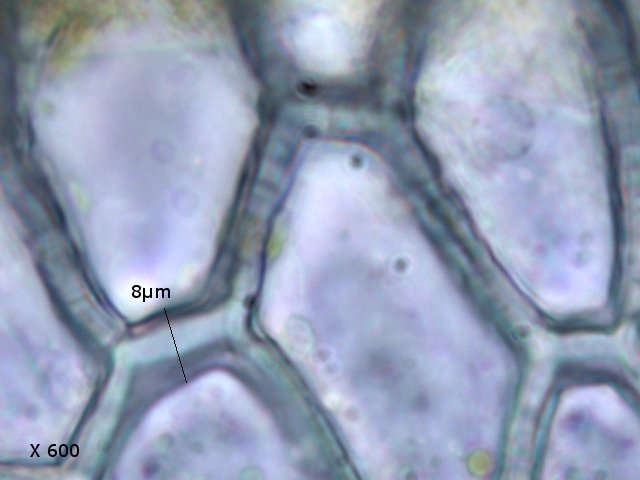
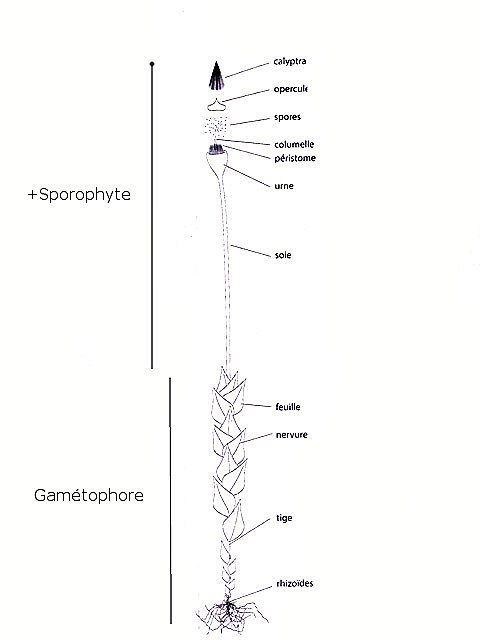
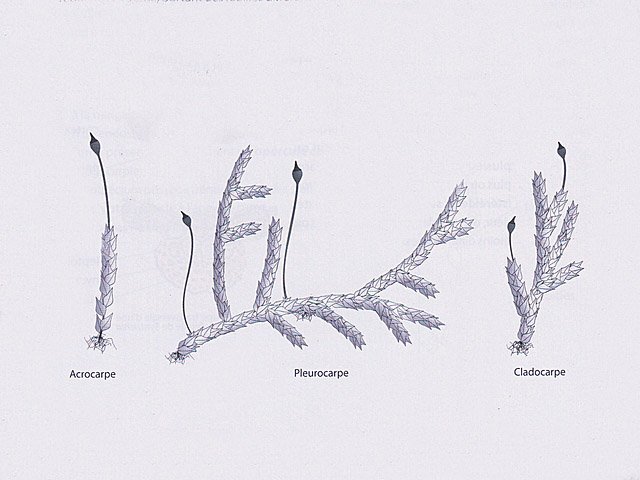
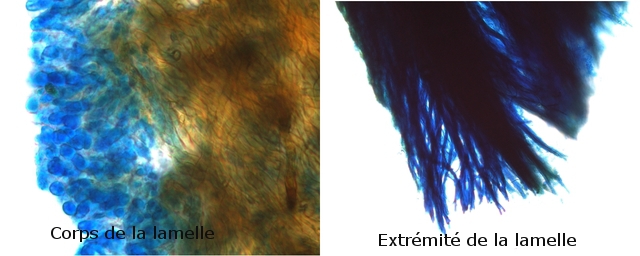
Mousse sur le toit Depuis plusieurs mois les mousses se développent sur le toit en tuile de la maison. Il semble y avoir plusieurs espèces .On va s’intéresser à l’espèce dominante Examen à la binoculaire A – Gamétophore B – Sporophyte Cette mousse se développe sur une hauteur de 2 à 3 cm suivant le lieu du toit et forme des plage de plus de 50 cm . Avant d’ aller plus loin considérons le schéma qui décrit le thalle des mousses Ce qui est caractéristique des mousses est que le Gametophore femelle ( qui est issu de la germination d’un Zygote ( 2n ) est porteur du Sporophyte - Il y a donc des gametophores mâles porteurs d’ anthéridies qui contiennent les spermatozoïdes flagellés ( absents de cette observation ) la fécondation des deux gamètes se fait dans l’ eau et donnera naissance à un prothalle puis à un thalle . La première constatation sur l’ échantillon est que le Gamétange a développé un rameau principal très ramifié avec des rameaux bien différentiés par rapport à la tige principale Les sporophytes se développant dans le prolongement de ces rameaux secondaires. Cette mousse est donc dite cladocarpe Les rhizoïdes : Les rhizoïdes servent à la fixation au sol Leur structure est pluricellulaire –les cloisons intercellulaires sont obliques La tige La tige est d’une largeur de 0,4 mm L’axe de la tige est occupé par un parenchyme médullaire ( il n’ y a pas ici de cylindre central ) On constate que les lamelles ( B ) se développent de part et d’ autres d’une fausse nervure ( A ) qui se fixe directement sur la tige ( il n’ y a pas de pétiole ) .Les feuilles ( lamelles ) sont développées sur deux axes opposés . La feuille est lancéolée, les bords sont lisses sans échancrure. Coupe d une feuille : La fausse nervure à fort grossissement : A – cellules internes à grand lumen -Leptoide qui sont des cellules conductrices B - et en periphéries les - cellules sont à lumière étroite à paroi épaissie --Stéréides qui sont des cellules de soutien. La feuille est sans nervure secondaire avec juste une fausse nervure centrale Les cellules du parenchyme des lamelles ( le limbe chez les feuilles angiospermes ) sont disposées en ligne allant du haut en bas de la feuille Sur la photo de gauche on découvre l’aspect du bord de la lamelle (feuille) Sur la photo de droite on constate que la paroi a développé des expansions sous forme de verrues dans lesquelles la cellule ne s’engage jamais Le Sporophyte ( voir schéma initial ): Le sporophyte est composé (voir schéma de départ) A Soie B Urne qui contient *** le peristome et la columelle qui ici ne sont pas visibles mais que l’ on devine avec la transparence ( première fermeture ) ***opercule qui ferme le contenu de l’urne (seconde fermeture) C la calyptra qui coiffe l’urne (troisième fermeture) Ainsi les spores qui sont contenues dans l’ urne doivent être libérées de 3 cloisons avant de pouvoir étre dispersées pas le vent Coupe du sporophyte à différents étages : Cette coupe du sporophyte a permis de mettre en évidence la présence de spores Elles sont mesurées ici autour 8 µm: La taille des spores des mousses est très variable - elle peut aller de 2 à 50 µm suivant les espèces Ces spores sont produites sur de longues périodes assurant ainsi un ensemencement efficace grâce au vent et à l’eau Quel est le nom de cette mousse ? La position cladocarpe des rameaux la fait classé dans le genre Dicranum Règne Plantae Classe Equisetopsida Sous-classe Bryidae Ordre Dicranales Famille Dicranaceae Genre Dicranum Par contre pour le nom binomiale les choses se compliquent Sur le site de MikrOscOpia il y a plusieurs Dicranum https://forum.MikrOscOpia.com/topic/5282-dicranum-viride-sull-lesq-lindb/#comment-22603 https://forum.MikrOscOpia.com/topic/598-dicranum-montanum/#comment-2394 https://forum.MikrOscOpia.com/topic/3834-dicranum-scoparium/#comment-18812 Les caractéristiques des mousses décrites dans ces observations ne collent pas bien avec l’ échantillon prélevé ici. Il a été décrit plus de 30 espèces ce qui nécessite des flores spécialisées Références Mousses et Hépatiques de France vincent Hugonnot et coll Edition biotope Biologie végétale Raven – Evert – Eichhorn Edition deboeck Dominique.
-
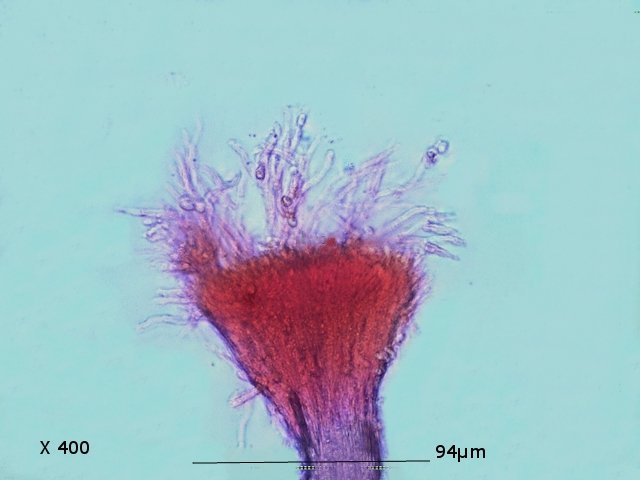
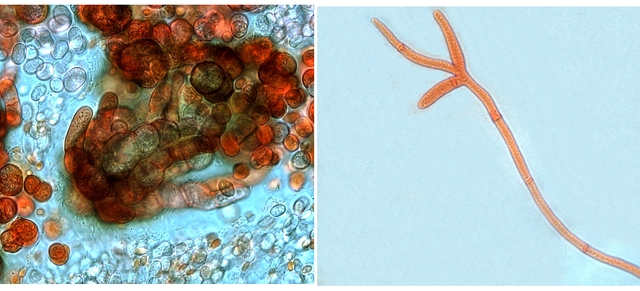
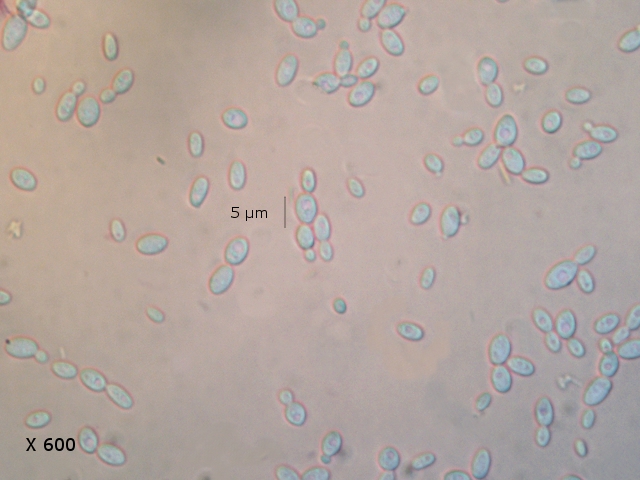
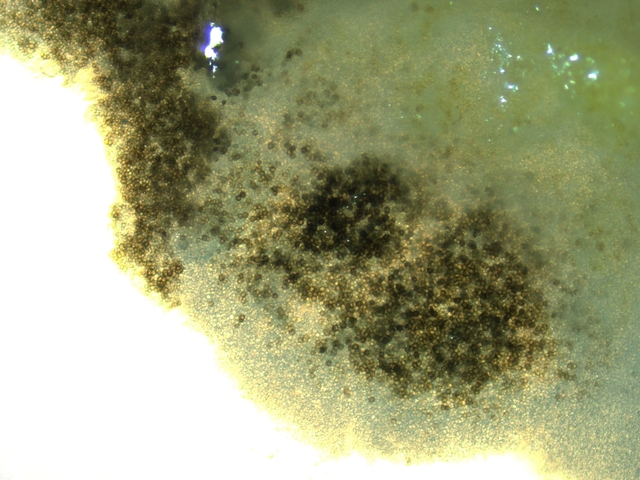
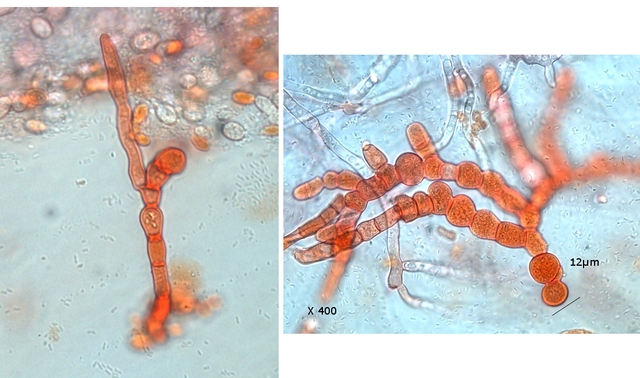
Graphium Moisissure du bois Planche En ce moment je fais des coffres pour cultiver mes fraisiers ; il est plus facile de les entretenir lorsqu’ils sont cultivés de cette manière et la production de fraises est excellente J’ai acheté des planches de coffrage chez le marchand du coin où les planches sont conservées au grand air protégées par un simple auvent La planche supérieure présente un enduit discret noir Les planches en dessous ne présentent rien comme particularité . A l’atelier sur la planche A il y a des trainées noires, rien sur la planche B qui a été protégée de ‘l’humidité de l’air . L’examen en episcopie d’ un copeau de cette planche montre : Le bois est couvert de nombreuses colonnettes noires comme des - poils dressés . Au sommet de quelques-unes on note la présence d’un petit point blanc. Exploration en episcopie à fort grossissement de cet échantillon. Aspect de têtes conidiennes Ces têtes ne sont pas rondes ni homogènes ( ce qui élimine un Rhizopus ) leur diamètre moyen est autour de 84µm. L’étude de cette moisissure va utiliser 2 colorants ( Le Bleu coton Lactophénol et Le rouge Congo ammoniaqué ). Image de la totalité du Thalle : Analyse de cette image : Les hyphes émergent du bois à certains endroits de la planche Elles prennent l’ aspect de rhizoïdes - En ( B ) les ponctuations des vaisseaux du Xylème des fibres de bois Surmontant ces hyphes s’est développée la tige : La colonette n’ est pas fait d’un seul hyphe mais de l’ association de nombreux hyphes fertiles ( A ) Ce regroupement se nomme une Corémie. Certains hyphes développent aussi des branches latérales ( B ) Wikipedia nous dit:Une corémie, ou synnema, est chez certains champignons une petite colonne non anguleuse composée d'un faisceau d'hyphes dressés et formant un conidiome, c'est-à-dire un regroupement de plusieurs conidiophores. Ce genre de structure est lié à la reproduction végétative car il produit des conidies. La corémie est dite « déterminée » lorsque la zone fertile est située dans la partie apicale de la colonne et « indéterminée » lorsque la zone fertile est située dans toutes les parties. Il existe aussi des corémies composées dont l'axe principal porte des ramifications latérales fertiles Ce qui est le cas de notre observation. avec un maximum apical donc cette moisissure développe une corémie de type déterminée La colonette est surmontée d’un buisson de conodiophores Ces têtes sont formées par le regroupement d’hyphes conidiogénes Les conidies ( spores ) arrondies elles semblent uniques au sommet d’un conidiophore et sont d’une taille autour de 2,30µm Cet aspect évoque beaucoup Le genre Graphium Domain: Eukaryota Kingdom: Fungi Division: Ascomycota Class: Sordariomycetes Order: Microascales Family: Microascaceae Genus: Graphium A noter que Graphium Ulmi -( autre nom Ophiostoma Ulmi) est un représentant de ce genre qui contient une centaine d’ éspéces Graphium Ulmi est responsable de la graphiose des Ormes et donc de la raréfaction de cet arbre https://atrium.lib.uoguelph.ca/items/cfedaa61-3d6f-4733-8592-78d38fab352e Référence https://drfungus.org/knowledge-base/graphium-species/ Graphium produces hyphae, conidiophores, synnemata, conidia, and rhizoid-like structures. Hyphae are septate and conidiophores are simple, long, and dark in color. Synemmata are bundles of erect hyphae and conidiogenous cells bearing conidia. The conidia are one-celled, oval, and colorless. They form clusters at the apex of each synnema. Rhizoid-like structures may be observed at the base of the synnema. Graphium spp. are recognized by their distinctive, erect, black synnemata, each bearing a single, terminal, ball of one-celled, hyaline conidia produced from annellides Dominique..
-
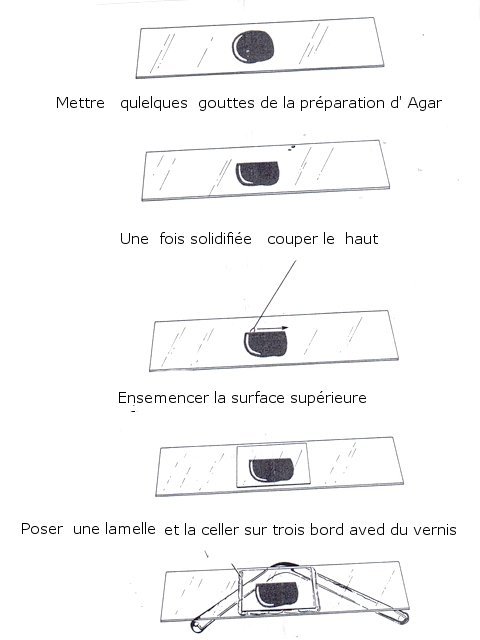
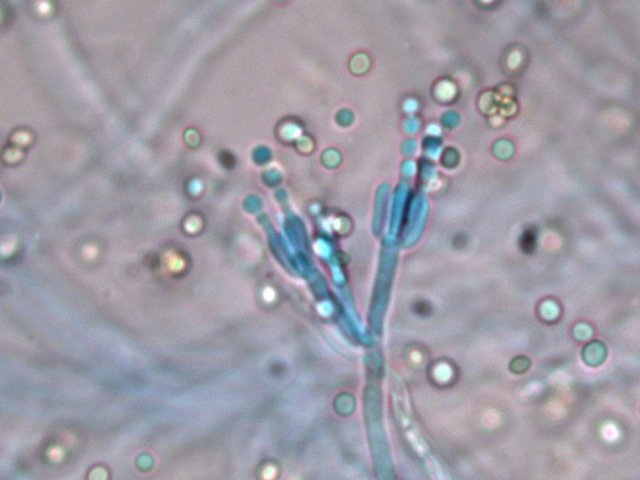
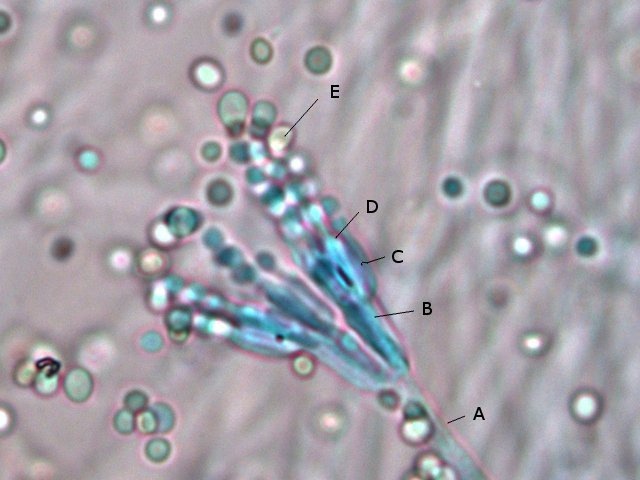
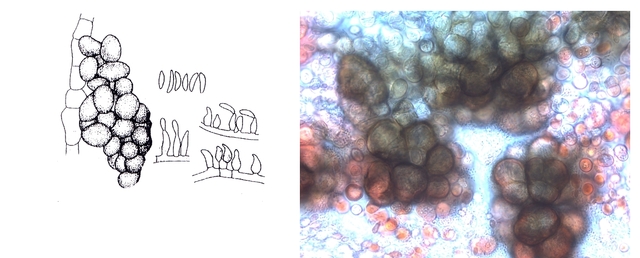
Moisissure mais laquelle ? Les moisissures nous entourent - il y en a dans tous les endroits où nous vivons. Et souvent dans les confitures ou les jus de fruits négligés dans une bouteille par exemple Mais comment savoir qu’elle espèce est présente. Si un prélèvement est fait à partir de la surface , une fois mis sur une lame il n’ y a que des spores et pas la moisissure reconnaissable avec le sporophore et ses conidies ou le sporange On ne reconnaît pas l’ organisation Le contenu de la bouteille est vidée sur un plateau. Une vue en episcopie souvent ne donne pas de bons résultats Il existe un moyen de faire mieux : Acheter de l’ Agar dans un magasin rayon pâtisserie Dans un verre Eau 1 cuiller à soupe 1/2 Agar une pointe de couteau ( 1 cm ) Sucre une pointe de couteau ( 0,5 cm ) Le mélange fond à 60 °c ( donc au bain-marie on constate que le milieu devient liquide ) A ce moment-là avec une petite tige on pose une goutte du mélange sur la lame - on ajoute goutte par goutte jusqu’ à obtenir le volume souhaité ( le mélange gélifie très vite ) couper le haut de la goutte - ensemencement mise d’un couvre objet – luter 3 côtés avec du vernis à ongles - conserver autour de 22°C Faisons l’expérience La lame est posée dans un boite de Pétri que l’on finit de préparer selon le schéma précédent - Aprés deux jours Examen au microscope Le problème est qu’ il existe une certaine distance entre lame et lamelle - l’ examen au microscope nécessite de varier la profondeur de champ . et qu’il faut un grossissement de 400. L’exploration de la lame est intéressante en raison du fait qu’ une très fine couche de gel se soit formée autour de la goutte.Cette zone est rapidement colonisée par les hyphes La première constatation est que les hyphes issus de spores différents ont tendance à s’ anastomoser X 400 A -- Conidiophore B Métule C – Phialide D --Formation des conidies sur la partie distale des phialides E – Conidies (spores ) Mais en Sciences rien n’ est simple , cette technique facile de mise en oeuvre donnera comme résultat la présence de Penicillium et de Rizopus ; Si on veut se faire développer d’ autres moisissures il faudra réaliser des mélanges nutritifs plus complexes ( ils sont nombreux et dépendent de la moisissure recherchée ). Vendus aux laboratoires par des fabricants - - Mais la technique utilisée pour la croissance de la moisissure est la même. milieu de Czapek milieu extrait de malt milieu à la farine d 'avoine etc .... Mais en Sciences rien n’ est simple , cette technique facile de mise en œuvre donnera comme résultat la présence de Penicillium et de Rizopus ; Si on veut se faire développer d’ autres moisissures il faudra réaliser des mélanges nutritifs plus complexes ( ils sont nombreux et dépendent de la moisissure recherchée ). Vendus aux laboratoires par des fabricants - - Mais la technique utilisée pour la croissance de la moisissure est la même. Références Moisissures utiles et nuisibles B Botton Masson1990 Détection et isolation des moisissures du sol P Davet INRA Dominique.
-
Lignification Topinambour Tige évolution au cours de l’année
Dominique. a posté un sujet dans Dominique
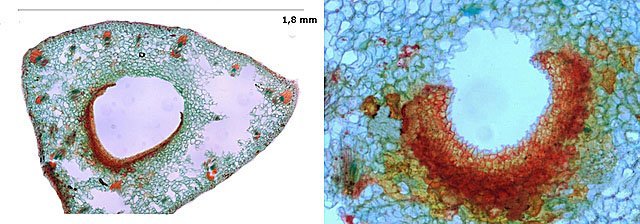
Lignification Topinambour Tige évolution au cours de l’année Le topinambour ( Helianthus tuberosus) est une plante de la famille des asteraceae bien connue Ces fleurs sont décoratives et ses tubercules sont comestibles et toujours présents sur les étals des marchés Sa croissance est rapide au printemps la tige est très souple -Cette tige depasse les 2 métres en automne et devient particuliement rigide - Comment expliquer cette rigidification ? Coupe faite le 12 Mai - (la tige atteint les 15 cm de haut ) A--Epiderme B –Parenchyme cortical C –Cylindre central D –Phloème E --Cambium F –Xylème G –Rayons médullaires H –Parenchyme medullaire Coupe faite le 16 juin Zone corticale A—Epiderme sur lequel sont apparus des trichomes (A’) B –Parenchyme cortical : son épaisseur a déjà beaucoup diminué. C –Cylindre central. Il se situe de plus en plus vers la périphérie au dépend de la zone corticale Axe vasculaire D –Phloème E –Cambium qui assure la croissance en centrifuge du Phloème et en centripète du Xylème Zone médullaire F –Rayons médullaires G –Parenchyme médullaire Coupe faite le 6 septembre A--Epiderme B –Parenchyme cortical : Axe vasculaire C –Phloème qui se lignifie D –Cambium E --Xylème F –On note la disparition progressive des rayons médullaires cellulosiques qui sont remplacés par l’apparition de faisceaux collatéraux lignifiés ce qui augmente la rigidité de la tige .On noter aussi le début de lignification des rayons medullaires qui restent ( couleur rosé ) F –Xylème Zone médullaire G –Parenchyme médullaire qui tend à disparaitre dans son centre Coupe faite le 5 Octobre (la tige atteint 1,90m) Zone corticale A—Epiderme Il s’est épaissi. B –Parenchyme cortical : Il a quasiment disparu. C –Cylindre central. Touche désormais l’écorce. Axe vasculaire D –Phloème .Il est totalement lignifié. E –Cambium Il s’est aussi lignifié (le diamètre de la tige est désormais fixé et cette lignification augmente la résistance de cette tige.) Zone médullaire F –On note la disparition des rayons médullaires cellulosiques qui sont remplacés par des faisceaux collatéraux lignifiés ). Conclusion La rigidité de la tige est liée au développement progressif de la lignification Ce processus de lignification débute très tôt dans l’année avec les vaisseaux de Xylème et s’ étend au Phloème puis aux cambiums . Par ailleurs en même temps que le parenchyme cortical se raréfiait sont apparus au cours de développement des faisceaux collatéraux qui se sont aussi lignifiés ( ces faisceaux collatéraux se sont développés au dépend des rayons médullaires cellulosiques qui sont de moindre résistance ). La lignification est donc un processus évolutif tout au long du cycle de développement de la tige du topinambour. Definition de Wikipedia La lignine (du latin lignum qui signifie bois) est une biomolécule, plus précisément une famille de macromolécules polyphénoliques, qui est un des principaux composants du bois avec la cellulose et les hémicelluloses. Ce biopolymère est présent principalement dans les plantes vasculaires et dans quelques algues rouges coralligènes, ce qui suggère une convergence évolutive de la biosynthèse des lignines entre ces algues et les trachéophytes. Ses principales fonctions sont de conférer de la rigidité et de la résistance mécanique aux parois cellulaires, ainsi que d'apporter une imperméabilité à l'eau et une résistance à la décomposition. **************************** Remarque technique sur cette observation : Le protocole de coloration a toujours été le même mais les résultats ne sont pas stables Pour les 4 préparations on a procédé de la manière suivante et pourtant les colorations obtenues sont vraiment différentes. Coupe autour de 120 µm Bain dans un mélange alcool/ acide acétique / formol 10 minutes ‘(AFA). Rinçage puis Eau de Javel du commerce durée jusqu’ au blanchiment des échantillons. Rinçage coloration avec un mélange (solution de safranine 3 gtt + eau 6 gtt + Solution d’Acridine 2 gtt ) 3 minutes Rinçage puis bain dans une solution d’Alun de Potasse 7 % 3 minutes Rinçage coloration avec le mélange (Astra Bleu 2 gtt + 6 gtt d’eau + Bleu Alcian 2 gtt ) 2 minutes Déshydratation avec un mélange Alcool 95 + Fotonal 1% (pour éviter les bulles d’air) Xylène quelques gouttes sur la lame (pour éviter les bulles d’air) Montage Euparal Explication possible un manque de rigueur dans les temps de manipulation des échantillons lors des différentes étapes ( et peut-être 1 goutte de plus ou de moins par mélange - mais la manipulation de la pipette Pasteur ‘n’est pas aussi évidente qu’ il y paraît ) . Dominique.
-
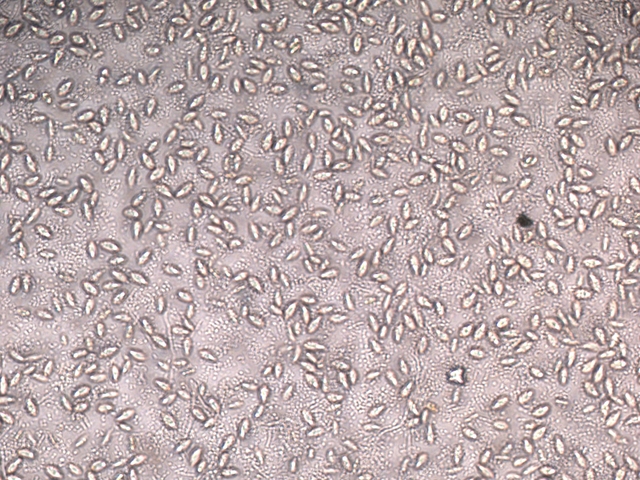
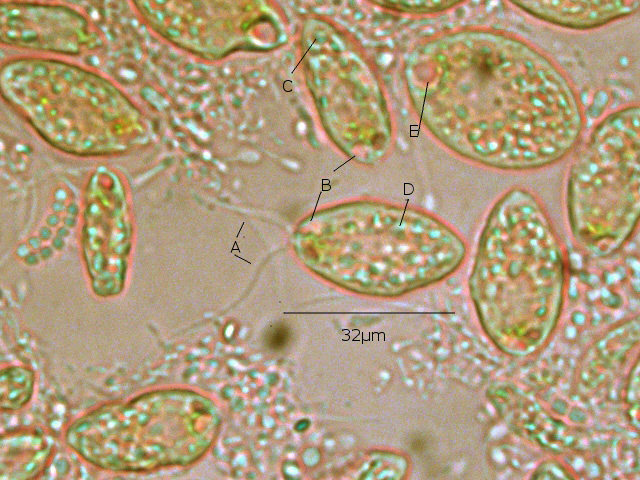
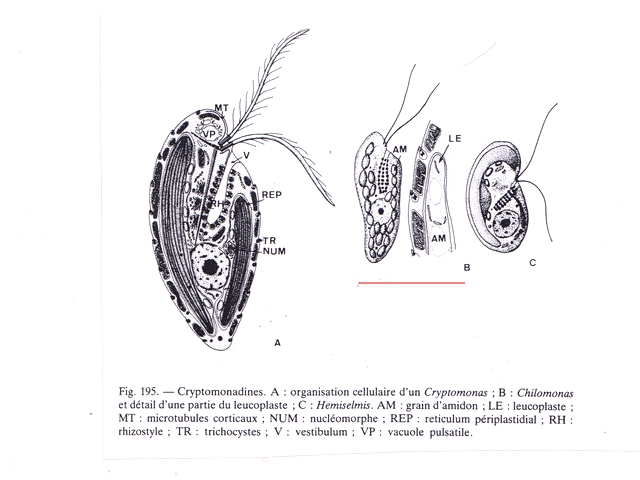
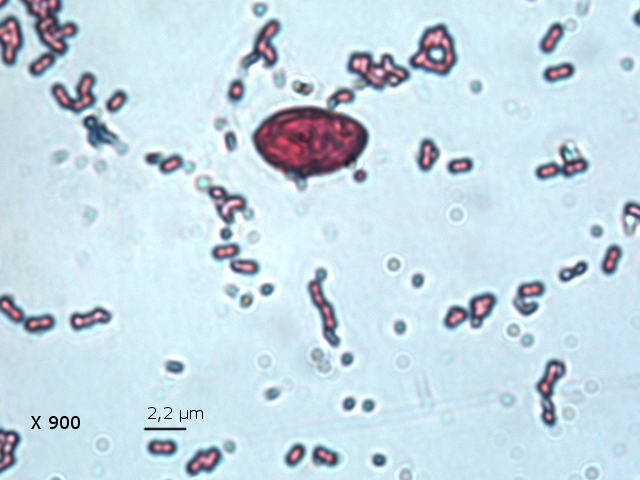
Film biologique Chilomonas Dans le jardin il y a un seau qui me sert à transporter les mauvaises herbes – la chaux - les fruits avariés et toutes sortes de choses Ce seau est resté sans emploi durant un certain temps - se remplissant doucement de l’ eau de pluie - ( mais pas depuis 15 jours car dans ce coin de Normandie la sécheresse s’ est installée ) Il y a quelques jours est apparu un film orange à sa surface. Ce milieu turpide révèle un Ph à 8,5 soit basique La face d’ ‘ une lame est posée sur la surface. Celle-ci est examinée au X 100 Sur cette photo on constate la présence de deux types d éléments dominants --A -la présence de protistes qui sont extrémement mobiles --B-- la présence de microbes Le protiste Examen au X 600 - ( le problème est que pour les forts grossissements tout ce petit monde bouge beaucoup ). Taille 32µm de long A - Ce protiste est porteur de deux flagelles à situation apicale B - A la base de ces flagelles il existe une dépression le cytopharynx C - Le noyau est situé dans la partie postérieure D Le cytoplasme est bourré de grains ( d’ amidon ?) E Il existe une vacuole contractile Cette description peut correspond au genre Chilomonas Dans le précis de Protistologie on peut lire « les plastes sont pariétaux de couleur souvent vert olive mais parfois brune , rouge ou même bleue » ce qui explique la coloration brune orangée du film ( l’eau en dessous du film est transparente ) . Classification: Empire: Eukaryota Kingdom: Chromista Subkingdom:Hacrobia Phylum Cryptista Subphylum: Rollomonadia Class: Cryptophyceae Order :Cryptomonadales Family: Cryptomonadaceae Genus :Chilomonas Les microbes Il est réalisé une coloration de Gram : technique violet de Gentiane 30 secondes - rincer - Lugol 20 secondes - rincer - Alcool 95 pour décolorer 5 à 10 secondes –Fuscine 30 secondes - Séchage à 40 °C Examen X 900 On obtient un résultat Gram négatif pour un microbe de type coccus associés par 2( Diplocoque ) . Conclusion : Les films biologiques sont en fait des milieux complexes - dans celui-ci il y a deux espèces dominantes Un diplocoque gram négatif et un protiste du genre Chilomonas . Il y a probablement d’ autres espèces mais elles ne sont pas facilement identifiables en raison du surpeuplement par la population de cocci et de protistes . Une autre observation avait déjà été faite de ce phénomène de film biologique avec pour peuplement dominant une algue https://forum.MikrOscOpia.com/topic/13864-algue-sur-une-mare/?hl=%2Bfilm+%2Bbiologique Références: https://eol.org/pages/49278345 Précis de protistologie de Pierre de Puytorac et coll edition Boubée 1997 – ( on prend conscience de la puissance des études qui ont été faite au 20éme siècle sur les protistes ( surtout en microscopie électronique ) – mais pour un amateur ce livre est illisible Dominique. dd
-
Pourrissement pêche agents responsables
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
Bonsoir Tryphon Je ne contredis pas - ton hypothèse en regard de la publication me semble seulement avoir une probabilité modeste . Mais cette situation est difficile devant l' accumulation de beaucoup d' imprécisions . . Amicalement Dominique -
Pourrissement pêche agents responsables
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
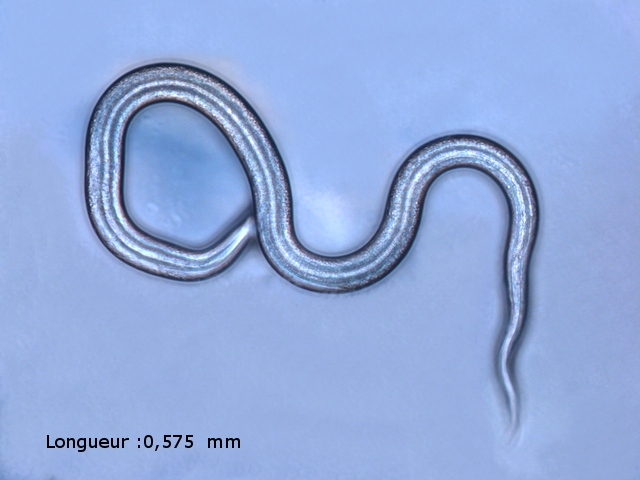
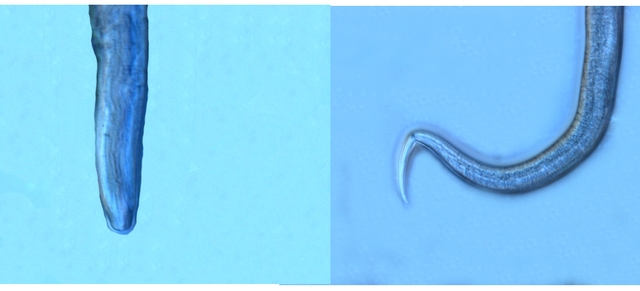
Bonsoir Jean -Marie et Tryphon Tryphon ton hypothèse est , en effet, correcte mais n' est pas acceptable si on se réfère au site suivant https://player.slideplayer.com/15/4736248/# il faut cliquer sur l'image pour les faire défiler Merci Jean - Marie pour l 'information . D'aprés cette publication on pourrait le classer parmi l' espéce Diphtherophora lata Amicalement Dominique -
Pourrissement pêche agents responsables
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
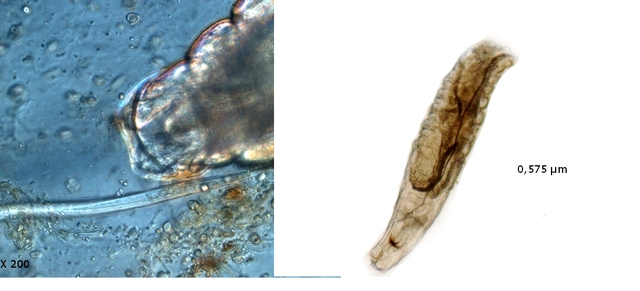
Bonsoir Jean - Marie Connais tu un site pour la reconnaissance des vers ? Je sais que l' image du pharynx est un élément important pour la détermination ( en agrandissant l' image on pourrait y trouver les indices nécessaires ) . Amicalement Dominique. -
Pourrissement pêche agents responsables
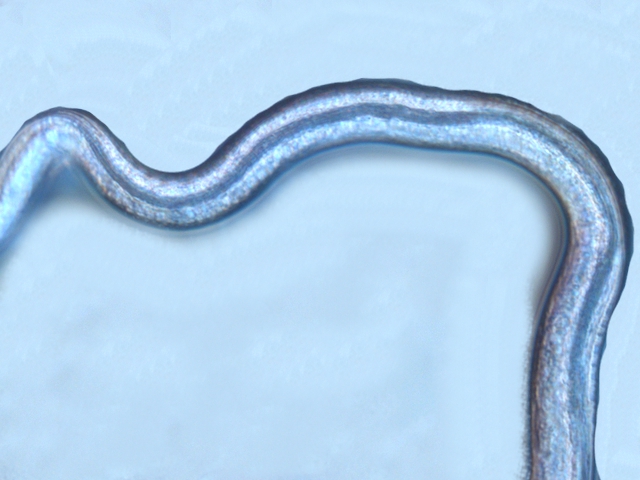
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
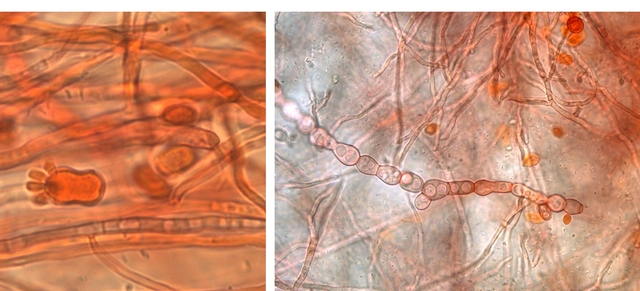
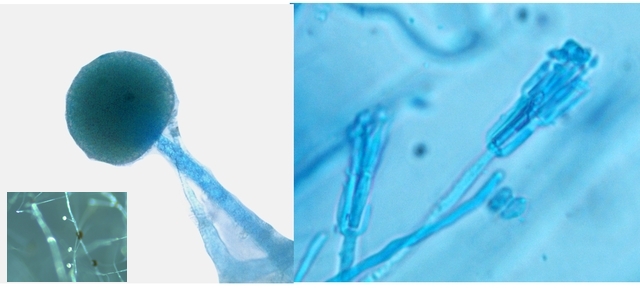
Les hyphes sont septés ( c’est à dire cloisonnés entre deux cellules fongiques ). On constate des cellules conidogénes ( production de spores directemement d 'une cellule d' hyphe . Ces aspects sont compatibles avec une moisissure du genre Microdochium . Cette moisissure est bien connue des cultivateurs puisqu’ elle fait partie des agents responsables de la Furasiose .Il s’agit d’ une maladie cryptogamique du blé . La fusariose des épis est provoquée par plusieurs espéces de fungus appartenant aux genres Microdochium et Fusarium L’infection entraîne souvent le blanchiment de tout ou partie de l’épi . La fusariose qui affecte l’épi peut entraîner des pertes de rendement pouvant aller de 30 à 70 %. Ces pertes sont dues à la nécrose précoce des épis et à des grains mal remplis et déformés; Une diminution de la qualité boulangère et du taux de germination. Les fusarioses sont à l’origine des mycotoxines. Prélèvement dans la partie profonde Ce prélèvement met en évidence la présence d’une grande quantité de vers Il existe un vers plat de plus grande taille qui se déplace sur la lame - il y en a très peu . sur la petite photo de droite on met en évidence la différence de taille entre les deux espèce - il y a une erreur sur le photo de droite : le vers plat fait 0,950 mm (soit une longueur un peu plus longue que le vers rond ) Je ‘n’ ai pas trouvé le nom de ces deux espèces . Les autres moisissures qui se sont développées sur le milieu de culture . A gauche - Sporange et Sporangiophore de Rhizopus ( en bas à gauche examen en episcopie ) A droite - Pénicillium Après 15 jours d’ évolution l’aspect de la pêche a changé - la couche de moisissures a disparu ou s’ est considérablement atténuée - par contre il existe une couverture grise feutrée dont on fait le prélèvement . Il s’agit d’ une colonie de Levures dont le développement a été permis par les modifications des tissus de la pomme sous l’effet de la moisissure et des bactéries . Enfin on constate comme à chaque fois qu’un fruit est laissé à pourrir au grand air le développement de drosophiles avec la présence à la base de la pêche d’un grand nombre de pupes https://forum.MikrOscOpia.com/topic/18780-trompe-mouche-drosophile/?hl=drosophile La pourriture d’un fruit devient un microcosme pour les saprophytes qui trouvent leur carbone dans la matière organique morte Ce monde est surtout dominé par les moisissures et les bactéries - J’ ai laissé le milieu évoluer - Après 15 jours le Penicillium et le Rizopus se sont installés en maîtres . puis ont quasi disparu pour être remplacés par une vaste colonie de levures - Après 15 jours les drosophiles sont toujours là. Ref Czapek https://fr.sperohope.com/agar-czapek-fundamento Fusariose du blé https://mycolab.pp.nchu.edu.tw/rice_fungi/species_info.php?species_name=Microdochium%20poae htt W. Microdochium trichocladiopsis (ex-type CBS 623.77). Scale bars: H = 100 μm; G = 50 μm; others = 10 μm.ps://plantpathogen.org/homepage/38-microdochium https://www.gene.affrc.go.jp/databases-micro_images_detail_en.php?id=30774 Dominique .
-
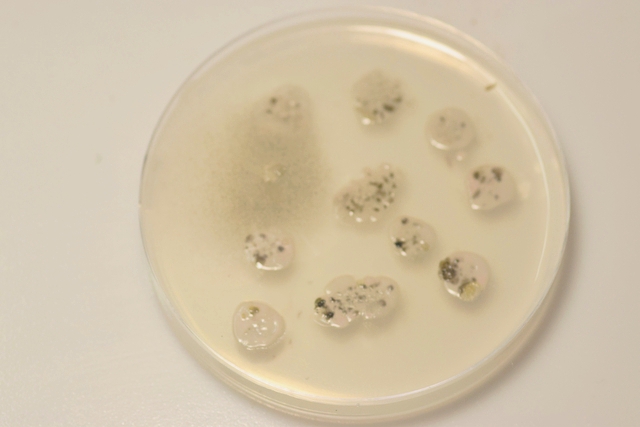
Pourrissement : le cas d’une pêche - les agents responsables Dans un coin du garage une pêche n’ a pas été emportée à la cuisine - Elle a subi son processus de dégradation habituel Mais quels sont les agents responsables de cette dégradation en dehors de l’inévitable prolifération bactérienne . Prélèvement de la couche superficielle : Le premier aperçu est celui d’une prolifération de moisissures - Les colonies bleutées sont habituelles sur les fruits en cours de pourrissement .Ce sont des colonies de Penicillium -mais quel est le nom de la colonie marron et noire ? . Un prélèvement local ne donne souvent pas de solution exploitable ( images microscopiques trop confuses ) d’où la nécessité de faire une culture .Un milieu de Czapek est fabriqué Les boîtes de Petri ensemencées avec un milieu dilué au 100 ( prélèvement d’un fragment mis dans un flacon de 10cc -prélèvement de 1 c dans 10 c - 2 fois de suite ) Étalement par gouttes sur le milieu et mise à l’ étuve à 25 °c :résultat 48 heures après Dans chaque goutte on constate le développement de colonies noires -A noter en haut à gauche une colonie de Rhizopus de croissance extrêmement rapide .et envahissante . Examen à la binoculaire de ces colonies noires : Prélèvement d’une colonie et étalement sur un lame - on fait une coloration au Rouge Congo Ammoniaqué. Le schéma initial résume les différents aspects que l’on va rencontrer en examinant la lame Sur la photo de droite il est mis en évidence ce qui ressemble à des sporodochies ( ce sont des regroupements d'un agrégat de filaments et de conidiophores le tout ayant une forme de coussin. Les cellules sont souvent groupées en stromas qui forment des masses irrégulières de filaments végétatifs.
-
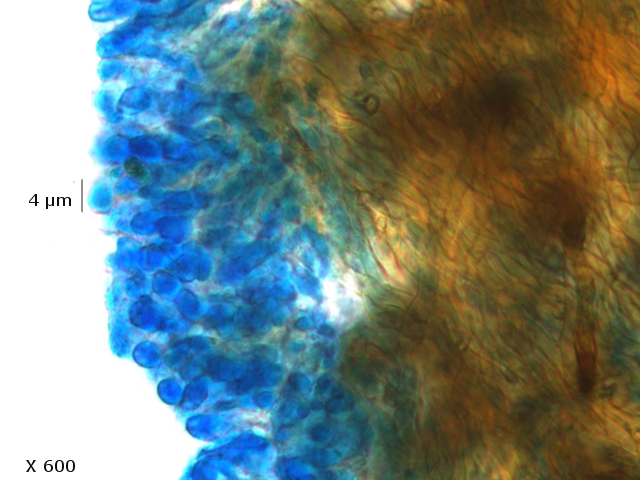
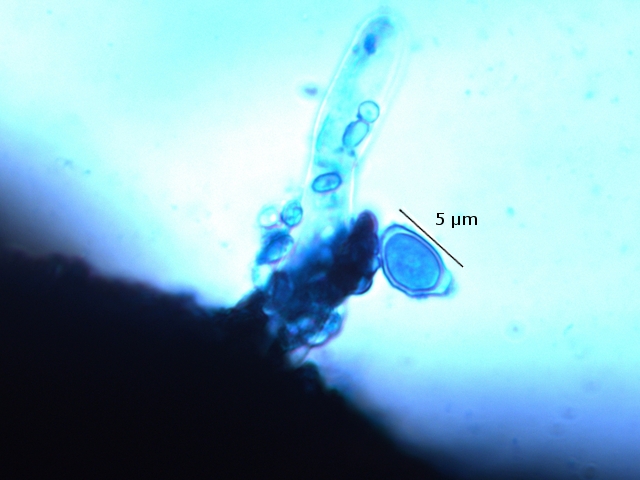
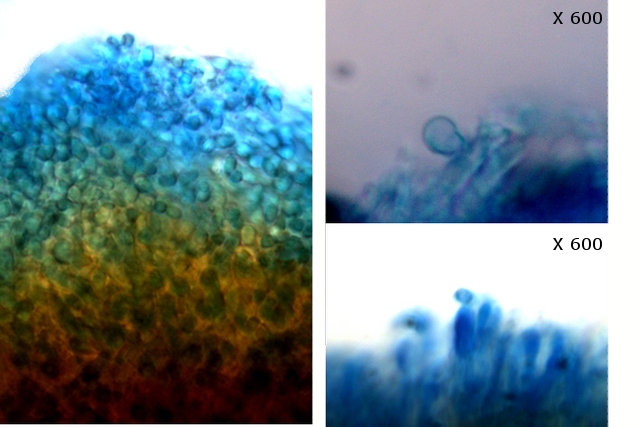
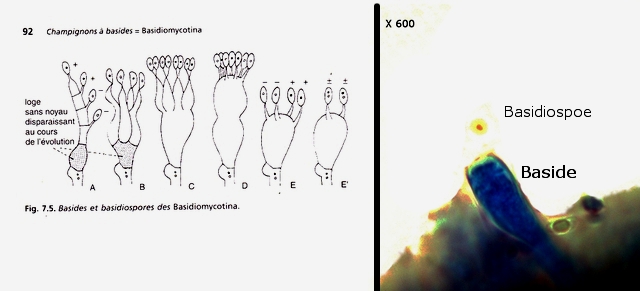
Polypore Genre Trichaptum Du fait de la sécheresse le sol de la forêt n’ est pas très riche en champignons - Les polypores par contre ne sont pas difficiles à trouver .Dans la rue du jardin botanique le tronc d’un vieux charme mort est toujours en place - Dessus existent plusieurs colonies de polypores . Ces colonies se développent depuis plusieurs années. Habituellement l'apparition des sporophores des champignons est intermittente du fait des variations climatiques et leur durée de vie est souvent très courte (quelques jours) ; mais le vie de certains sporophores peur dépasser quinze ans chez les polypores coriaces comme c’est le cas pour le genre Trichaptum Cette longue survie implique une structure qui puisse résister aux intempéries. Exploration de la structure de ce Basodiomycéte . Coupe d’une langue de ce polypore La coupe met donc en évidence 3 zones Une zone d’ attache au tronc - d’où partent les hyphes qui vont coloniser le bois en cours de décomposition - Une zone épaisse qui assure la rigidité et la fixation Une zone fertile périphérique , située en position inferieure, où vont se développer le sporocystes Une zone d’ attache au tronc - d’où partent les hyphes qui vont coloniser le bois en cours de décomposition - Une zone épaisse qui assure la rigidité et la fixation ( X100 ). Partie superieure : Cette zone de coupe est située près de sa fixation sur l’ arbre – il est constaté que les hyphes semblent morts et qu’ils forment un amas compact du moins dans sa partie supérieure. Coupe en dessous de la zone ci-dessus qui est la zone de circulation entre les hyphes qui pénètrent le bois et le sporophore La structure est formée d’un emmêlement d’un grand nombre d’ hyphes. La zone génératrice :Les lamelles sont situées sur la partie inferieure du sporophore : Vue de la face inferieure sur un sujet plus jeune que celui utilisé pour les coupes précédentes. Coupe d’une lamelle X 100 Les zone fertiles sont donc situées de part et d’autre de la lamelle qui est formée par un compactage d’ hyphes : Images précisant le développement reproducteur et la zone inter lamellaire dans sa partie distale . Les spores : L’examen des coupes met aussi en évidence trois types de spores : Les conidies ( ou spore asexuées Le plan profond est formé par de très nombreux hyphes - Le plan superficiel est lui formé de structures cellulaires rondes /ovalaires plus ou moins régulièrement empilées . devant correspondre à des empilements de conidies (Une conidie (ou conidiospore) est une spore assurant la multiplication asexuée des champignons-). Les chlamydospores ( spore asexuée de résistance ). Les chlamydospores sont des spores asexuées de forme sphérique ou ellipsoïde, produites par certains champignons basidomycètes. Elles sont généralement plus grandes que les spores sexuelles (basidiospores) et sont entourées d'une paroi épaisse et résistante. Les chlamydospores sont produites dans diverses conditions, notamment en réponse à des conditions défavorables, telles que la sécheresse, le froid ou la malnutrition. Elles permettent au champignon de survivre à ces conditions et de se propager lorsque les conditions redeviennent favorables. Les basidiospores . Dans cette observation malgré un nombre important de coupes il a été très difficile de mettre en évidence des basides et encore moins des basides correctes surmontées par son image bien connue avec ses 4 spores .Cette forme sexuée de reproduction n’ est donc pas la forme dominante . Ces spores sont en effet la forme sexuée de la propagation du champignon . Ces spores sont en effet la forme sexuée de la propagation du champignon . La baside sur la droite de la photo est de qualité plus que modeste , elle est porteuse d’une seule basidiospore et le stèrigmate qui la lie à la baside n’est pas visible . Les basidiospores sont formés par l’union des deux noyaux haploïdes ( n chromosomes ) des cellules d'hyphes de polarités différentes ( on ne parle pas de sexe ) qui convergent deux à deux, ces deux noyaux coexistent d’abord sans caryogamie puis s’unissant forment un Zygote à 2n chromosomes Ce zygote va subir une méiose qui va donner naissance à 2 noyaux haploïdes ( n chromosomes ) En fonction du nombre de mitoses qui vont suivre on va obtenir 2 ou 8 spores - ( mais beaucoup ne se développeront pas ). Kingdom: Fungi Division: Basidiomycota Class: Agaricomycetes Order: Hymenochaetales Family: Incertae sedis Genus: Trichaptum Murrill (1904) Référence : Les champignons mycologie fondamentale et appliquée de P Boucher et col editeur Masson Dominique
-
Bonsoir Jean -Luc , Jean-Marie , tous. Les coupes ont été faites après avoir placé l' élytre sans préparation dans un moule où j' ai coulé de la paraffine - le bloc a été coupé au Microtome de Minot 70 µm d' épaisseur- Le mécanisme d' ouverture des élytres est musculaire seul l' aile s 'ouvre par pression hydraulique ( hemolymphatique ). Amicalement Dominique.
-
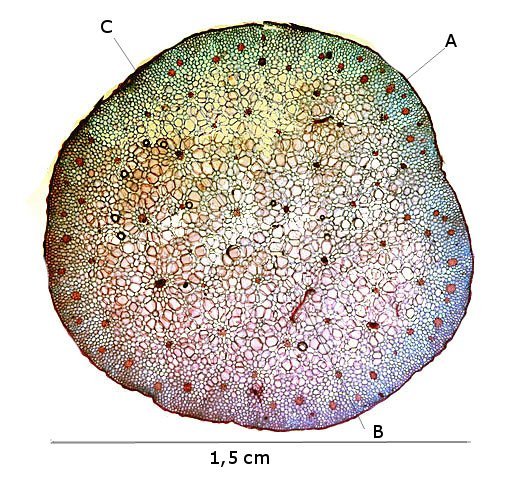
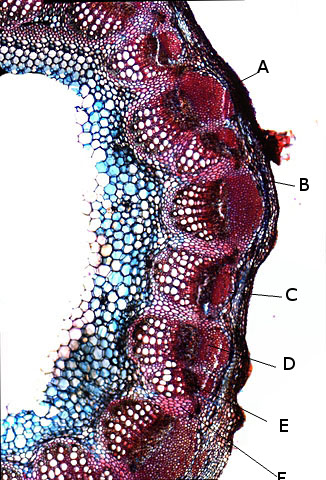
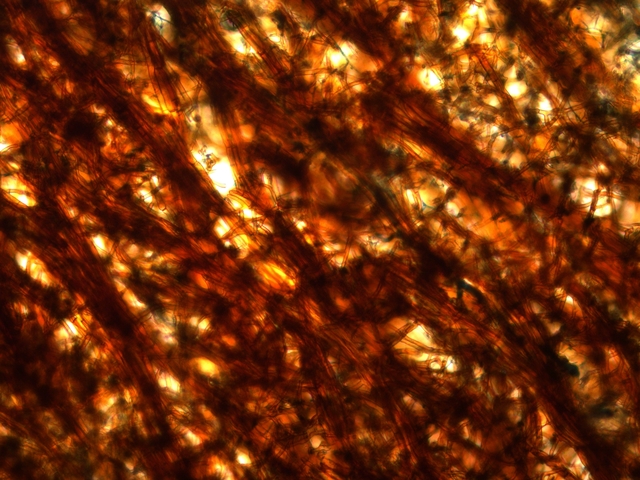
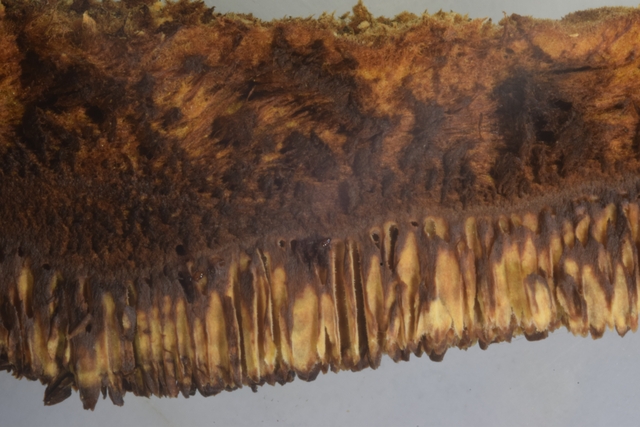
Coléoptère élytres ailes Lors d’une marche dans la forêt de Balleroy ( 14 ) , on découvre , gisant sur le sol une lucane cerf-volant femelle - avec une élytre entre ouverte . Les coléoptères ont deux paires d’ ailes - 2 ailes du plan supérieur ou élytres - et 2 ailes du plan inférieur membraneuses . Au toucher l’élytre est rigide - franchement dur - non pliable .Une coupe de cette élite est réalisée : La résistance de cet organe tient à sa structure - Il est constitué de plaques comme sont réalisées les poutres en lamellé-collé dont la résistance est telle qu’ elles permettent des portées de 45 mètres ; Image en lumière polarisée . L’aspect est celui d’un empilement cellulaire qui s’organisent par plaques . Les élytres sont constituées - comme le reste du squelette externe des insectes de chitine - Cette protéine est renforcée par la présence de calcium et de phosphore L'élytre des coléoptères est constituée de trois couches cellulaires principales : -- L'épicuticule est la couche externe de l'élytre. Elle est composée de chitine et de lipides. Elle est imperméable à l'eau et à l'air. --L'endocuticule est la couche intermédiaire de l'élytre. Elle est composée de chitine et de protéines. Elle est plus flexible que l'épicuticule. --L'hypoderme est la couche interne de l'élytre. Elle est composée de cellules vivantes. Ces cellules produisent la chitine et les autres substances qui constituent l'élytre. En plus de ces trois couches cellulaires principales, l'élytre contient également des poils, des glandes et des cellules sensorielles Les glandes sont présentes dans l'hypoderme. Elles produisent des substances telles que de la cire, des phéromones et des toxines. Les élytres protègent deux ailes membraneuses : Ces ailes membraneuses ont 2 positions : Repliées : Ouvertes : Comment ces deux ailes pliées peuvent elles se déployer ? ( elles le font avec une très grande rapidité comme on peut facilement le constater quand on assiste à l‘ envol d’une coccinelle ). L’aile est pourvue de deux articulations : Articulation sur le bord d’attaque ( X 40 + recadrage). Articulation au milieu de l’ aile sur une grosse nervure ( X 40 +recadrage ) Par quelle mécanisme ce déploiement des ailes peut-il se faire Puisque il n’est pas possible de mettre en évidence la présence ni de muscle , ni de tendon ? En réalité l’examen bord des nervures semble apporter la solution : Articulation antérieure : photo prise le long du bord d’attaque : Articulation du milieu de l’ aile le long d’une nervure : Sur ces deux photos au X 100 il est mis en évidence la présence d’un système vasculaire - Le moteur du mouvement des ailes est donc de type hydraulique à l’ image des bras des gros instruments de travaux publiques - L’ hémolymphe comprimé par les muscles thoraciques va conduire à une modification de la forme des articulations dont la conséquence sera l’ ouverture de l’ aile ( ce qui explique que les articulations ne sont pas comme celle de notre fémur avec le bassin ou de notre humérus avec l’ omoplate qui pour fonctionner nécessité la traction de tendons ) Ici c’est la déformation des surfaces articulaires qui va créer le mouvement . Le mouvement des élytres par contre se déplie pas activité des muscles et des tendons thoraciques . Régne Animalia Embranchement Arthropoda Sous-embr. Hexapoda Classe Insecta Sous classe Pterygota Infra classe Neoptera Super ordre Endopterygota Ordre Coleoptera Sous-ordre Polyphaga Infra-ordre Scarabeiformia Famille Lucanidae Genre Lucanus Espéce Lucanus cervus Quasi menacé Dominique
-
Bonjour Jean-Marie Je pense que l’ on peut placer ces plantes dans un jardin public à condition de les enlever dès que les fruits sont mûres .Cela nécessite donc une surveillance Je vais faire attention à l’ évolution de ces plants -( sinon je signalerai à la municipalité le risque et la nécessité de les enlever dès la fin aout - période ou les graines ne sont pas encore expulsées ) - - Ces plants ont-ils une toxicité moindre ? - il faudra se renseigner . Amicalement Dominique
-
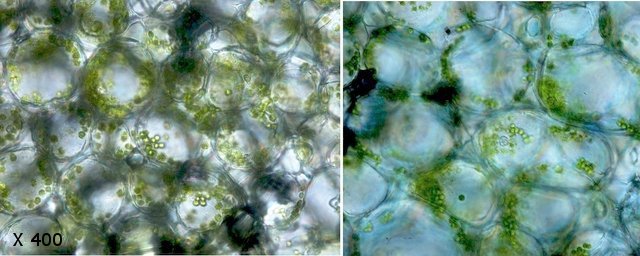
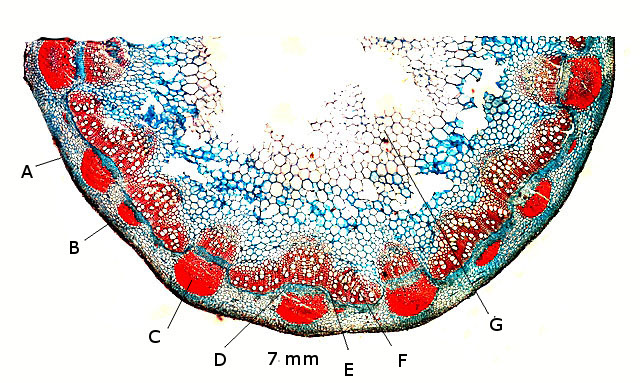
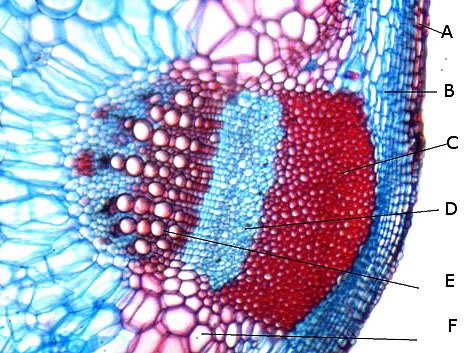
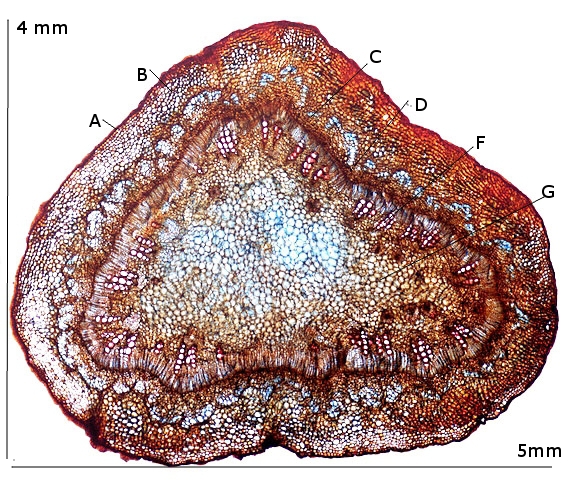
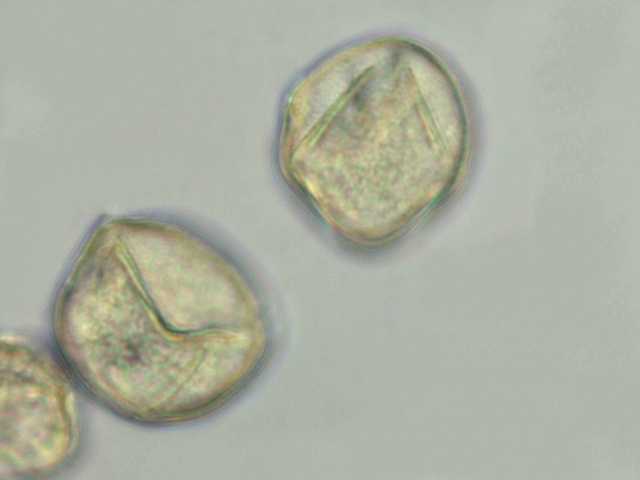
Ricin commun Ricinus communis Cette année la municipalité a décidé de placer des plants de Ricin sur les massifs du parc public-le résultat est très agréable :cette plante par son feuillage et l’ aspect inhabituel de ses feuilles donne une impression d’ exotisme dans ce petit port normand . Les fleurs sont regroupées ( Formant des cyathes - caracteristiques des euphorbiacés dont fait partie le Ricin commun Les fleurs mâles et les fleurs femelles sont portées par la même plante ( La plante est dite monoïque). Elle fleurit avec le début de l’ été . Je propose de regarder cette plante au travers du microscope La feuille La tige La fleur mâle La fleur femelle - la graine La feuille Les feuilles sont palmées (avec 5 à 7 lobes pointus), dentelées, vertes avec des nervures plus ou moins rouges. Elles sont reliées à la tige principale par de longs pétioles (30 centimètres). Coupe de la feuille à hauteur d’une nervure A – Epiderme B – Periderme (Phelloderme -phellogène) C – Zone qui contient de la périphérie vers le centre : les cellules de collenchyme – les cellules parenchymateuses - le cellules de sclérenchyme D – Xyléme primaire D – Xyléme secondaire E – Cambium Coupe d’une feuille à hauteur du limbe A –Cuticule B --Cellules palissadiques C –Cellules du mésophile D –Lacunes aériennes ( contenant l’ oxygène produit par le processus de photosynthèse et le CO2 d’origine extérieur ) E –Epiderme inferieur La tige principale A – Epiderme B – Periderme (assise subéro-phellodermique ou cambium subéro-phellodermique, est un méristème secondaire cortical ) C –,Endoderme correspond à l’extrémité du cylindre central d’un faisceau libéro-ligneux– Phloeme secondaire D – Cambium périphérique F – Xyléme G –Cellules médullaires La tige porte des fleurs mâles et femelles A— Fleur mâle ouverte avec son bouquet d’étamines. B -- La fleur femelle a ici laissé la place aux fruits en cours de développement. Les fleurs mâles sont groupées le long de la tige : L’ efflorescence des petites fleurs mâles , (Cyathe de fleurs mâles )- Par contre dès leur ouverture elle développent des bouquets blancs Ces étamines ont la particularité de former des arborescences par division dichotomique des filets ; L’anthère est fixée à chaque fin de division dufilet de l'étamine par le « connectif », Elle est composée de deux thèques possédant chacun deux sacs polliniques avant maturité, nombre constant chez les Angiospermes. Ceux-ci fusionnent à maturité pour donner deux loges (anthère biloculaire),. Elles s'ouvrent, généralement par déhiscence, pour répandre les grains de pollen à l'extérieur. L’organisation des étamines: Les anthères: La dispersion se fait par le vent . Le pollen: Taille des pollens autour de 50 µm . Les fleurs femelles Elles possèdent 5 tépales Elles forment des inflorescences très rouges. La fleur du ricin commun a 3 carpelles. Fusionnés qui forment le Gynécée. L’ensemble des carpelles forme le pistil central surmonté de 3 stigmates qui sont bifides L'ovaire contient 3 ovules, Photo des 3 stigmates : Les fruits Ce sont des capsules tricoques : Ces fruits sont l’un des éléments les plus spectaculaires de la plante .Leur péricarpe rouge est très ornementé de longues pointes . La coupe d’une graine : Ces graines ont des propriétés toxiques, La toxine : la ricine est capable d’inhiber la synthèse des protéines en attaquant l'ARN des ribosomes, entraînant la mort cellulaire. Elle est présente à une concentration variant de 1 à 10 % dans la graine de ricin. Il est réalisé une coupe du péricarpe qui enveloppe la graine : A -- Téguments de la graine. B -- L'endocarpe est la couche la plus interne du péricarpe. Il est généralement mince et adhère à la graine C – Le mésocarpe est la couche intermédiaire du péricarpe. Il est généralement plus épais que l'épicarpe D – L'épicarpe est la couche la plus externe du péricarpe. C’est cette couche qui donne cet aspect spectaculaire à ce fruit grâce au développement d’ importantes ornementations . La graine : Les graines seront évacuées par des déhiscences latérales qui au mois d’ Aout ne sont pas encore visibles - Classification classique Règne Plantae Classe Magnoliopsida Ordre Euphorbiales Famille Euphorbiaceae Genre Ricinus Nom binominal Ricinus communis L., 1753 Pour en savoir plus : https://fr.wikipedia.org/wiki/Ricin_commun La totalité de la plante semble toxique en raison de la présence d'une lectine glycoprotéique : la ricine. La concentration en ricine est maximale dans les graines qui renferment par ailleurs des protéines, de l'eau et des lipides. Ces graines sont riches en une huile qui doit ses propriétés purgatives à la présence de l'acide ricinoléique. qui altère la muqueuse intestinale et provoque des pertes importantes en eau et en électrolytes (sels minéraux), d'où son action purgative intense et irritante. La ricine, protéine présente dans la plante et les graines, est une toxine . . Le passage à l'autoclave de la farine pendant 15 minutes à 125 °C détruit la ricine. On considère que trois graines peuvent être fatales à un enfant, quatre graines peuvent déterminer une intoxication sérieuse chez l'adulte et six à huit graines pourront lui être fatales. Les pigeons ramiers sont également sensibles aux graines de ricin et de nombreux cas d'intoxication ont été constatés dans les villes qui en utilisaient comme plante ornementale. La production des graines de ricin dépasse les 100 millions de tonnes par an ( Production surtout en Inde - en Chine et au Brésil ) On l'utilise depuis peu comme source de biocarburant. L'huile est également utilisée comme lubrifiant dans les moteurs de voitures de course et de modèles réduits. Il entre dans la composition d'une matière plastique (non-biodégradable) nommée Rilsan. Dominique.