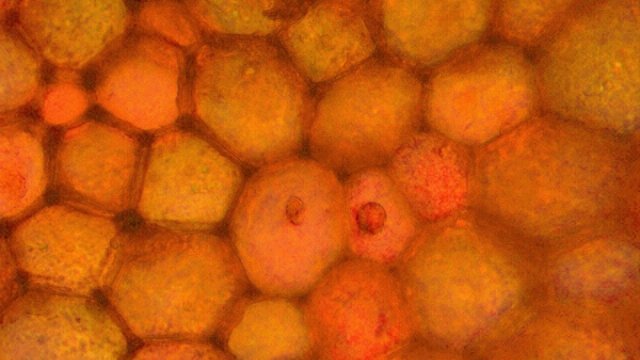Dominique.
-
Compteur de contenus
1 481 -
Inscription
-
Dernière visite
-
Jours gagnés
1
Tout ce qui a été posté par Dominique.
-

Petite collection de copépodes marins
Dominique. a répondu à un(e) sujet de Jean Marie Cavanihac dans Jean Marie CAVANIHAC
Bonsoir Jean -Marie Existe t il des différences qui permettent de dire ce copépode est d'origine eau douce celui-là vient de la mer ? Ta dernière photo est particulièrement réussie . Amicalement Dominique -
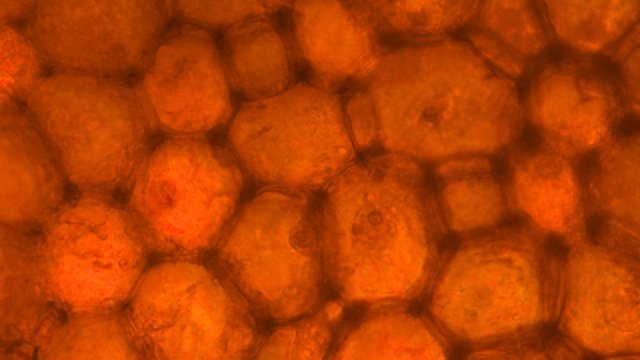
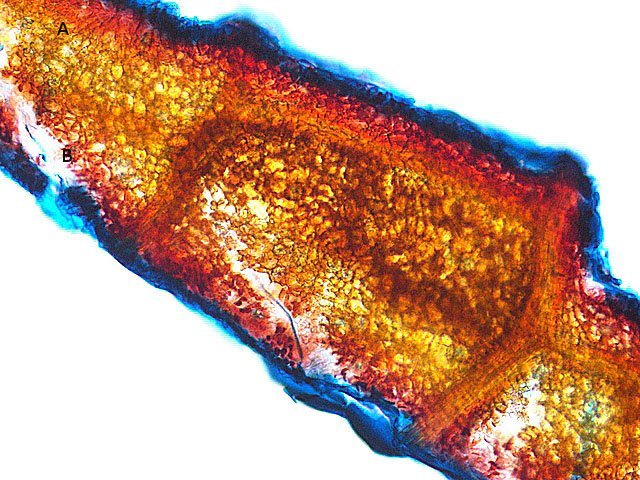
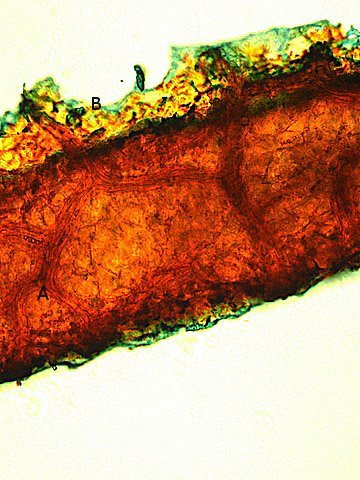
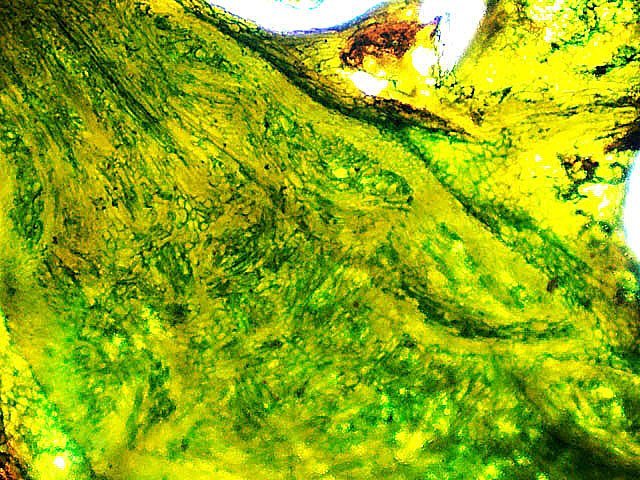
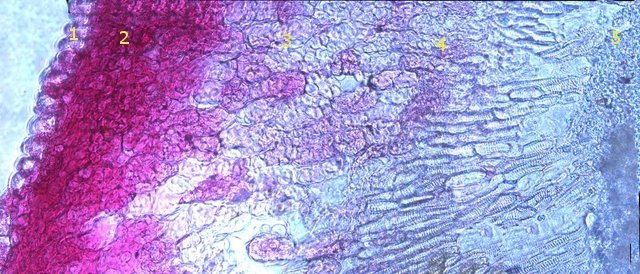
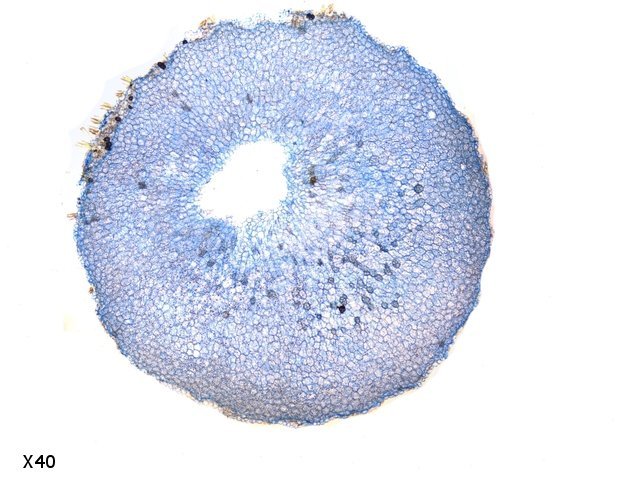
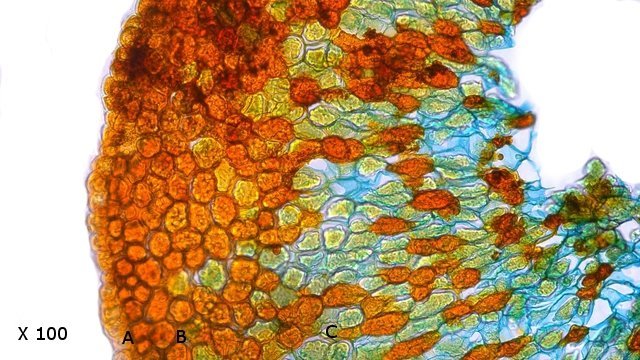
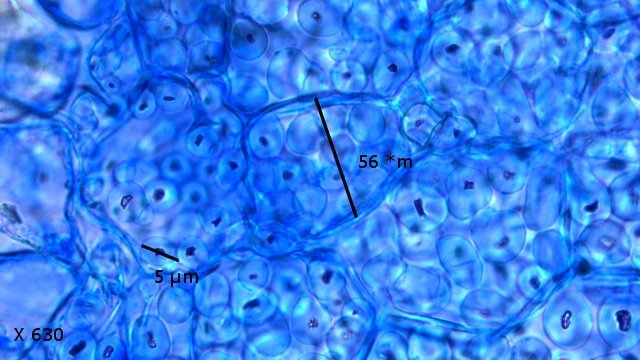
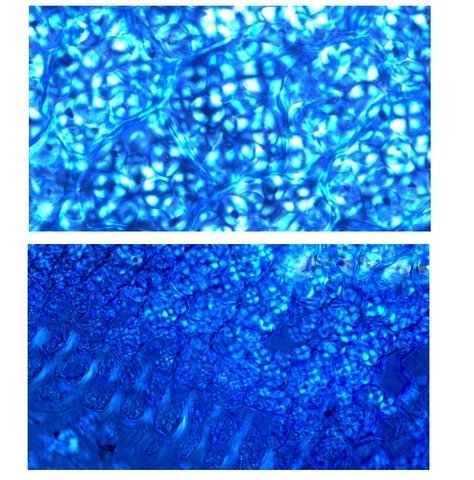
Galles sur feuille de chêne suite Dans le premier article qui a abordé la galle du chêne https://forum.MikrOscOpia.com/topic/20210-discomyc%C3%A8tes-sur-feuille-de-ch%C3%AAne/ Jmaffert a redressé un diagnostic erroné et Tryphon nous a fourni un lien très intéressant sur les galles https://www.zoom-nature.fr/la-galle-lenticulaire-du-chene/ Dans ce second article l’ intérêt est porté sur la structure des galles En effet sur la même feuille il a été possible de découvrir 4 galles différentes 3 déjà présentées. Photo N°1 galle suite à la piqure de la guêpe ? Photo N° 2 galle suite à la piqure de la guêpe Neuroterus quercusbaccarum Photo N°3 galle suite à la piqure de la guêpe? ( sur le Web il apparaît une grande confusion dans les dénominations des guêpes) Et une quatrième pas encore photographiée. : Il s’ agit d’une galle suite à la piqure de la guêpe :Neuroterus numismalise . En coupe : Des coupes sont réalisées Coupe après inclusion dans la paraffine section de 30 µm. Coloration avec Acridine /Acriflavine/Bleu Alcian . Bleu Alican seul Bleu coton lactophénol . D’abord deux coupes sans aucune préparation ni coloration : 1 1 Cellules épithéliales 2 2 Cellules colorées naturellement en rouge indice de la présence de caroténoïdes 3 Parenchyme 4 Réseau vasculaire 5 Cellules limitant la cavité de développement de la larve Coupe colorée au Bleu Alcian après la préparation classique alcool/acide acétique / formol . Coupe colorée après préparation Acriflavine/Acridine/Bleu Alcian A-cellules épithéliales B-cellules lignifiées C-cellules à paroi cellulosiques Réalisation de coupe de la Galle N°1: A – zone d’attache à la feuille, lieu où passent les vaisseaux reliant le système circulatoire de la feuille au système circulatoire de la galle. B --loge où la larve peut se développer. C –parenchyme fait surtout de cellules à paroi cellulosique riche en plastes D –parenchyme périphérique riche en cellules en lignifiée assurant la solidité de la structure. Les coupes suivantes ont été colorées avec Le bleu Coton lactophénol : En moyenne les cellules mesurent autour de 56 µm Les plastes mesurent autour de 5 µm Si on utilise la lumière polarisée on retrouve la croix caractéristique créée par la présence de l’amidon Les plastes sont donc des amyloplastes. Coloration avec du Lugol ( iode ): Une grande quantité de cellules prennent le colorant confirmant la présence de l’amidon. Dernière image sur la périphérie de la galle N°4: Les cellules épithéliales sont couvertes (A) par une épaisse couche de cire isolante ( cuticule ). ****************************************************************************************************************** Commentaire sur la formation des galles (Cécidogenése ): La formation d’une galle après une piqûre de guêpe (ou plus souvent d’un insecte comme un cynips, un puceron ou un acarien) est un phénomène qui relève d’une interaction complexe entre la plante et l’insecte.La larve qui se développe dans la galle semble aussi synthétiser des molécules actives sur la croissance de la Galle. Voici comment cela s’explique : 1. Une croissance cellulaire contrôlée, pas anarchique La galle n’est pas une simple prolifération désordonnée de cellules. Elle résulte d’une reprogrammation ciblée du développement de la plante par l’insecte. Les substances injectées (salive, toxines, hormones) agissent comme des signaux qui modifient l’expression de gènes spécifiques de la plante, notamment ceux impliqués dans : La morphogenèse (forme de la galle) La différenciation cellulaire (types de cellules formées) La division cellulaire (où et comment les cellules se multiplient) Cette interaction est si précise que chaque espèce d’insecte produit une galle avec une architecture caractéristique (taille, forme, texture, couleur), presque comme une "signature". Ces différences reflètent des mécanismes moléculaires distincts, où l’insecte "reprogramme" la plante de manière spécifique. 4. Preuves scientifiques récentes Des études en génomique et en biologie moléculaire ont montré que : Les insectes gallicoles injectent des ARN ou des protéines capables de modifier l’expression des gènes de la plante hôte. Certaines galles expriment des gènes normalement actifs uniquement dans des organes spécifiques de la plante (comme les racines ou les fleurs), ce qui suggère une redirection du programme de développement. 1. L’injection de signaux par l’insecte Lors de la piqûre ou de la ponte, l’insecte injecte un cocktail de molécules dans le tissu végétal. Ces molécules incluent : a. Effecteurs protéiques Fonction : Ces protéines, souvent sécrétées par les glandes salivaires de l’insecte, ciblent des voies de signalisation spécifiques de la plante. Protéines similaires à des hormones végétales (comme les auxines ou les cytokinines) qui mimétisent ou perturbent les signaux de croissance de la plante. Enzymes modifiant les parois cellulaires (pectinases, cellulases) pour faciliter la pénétration et la diffusion des signaux. Protéines inhibitrices de défenses (comme des inhibiteurs de protéases ou de l’éthylène) pour supprimer les réactions immunitaires de la plante. b. MicroARN (miARN) et ARN interférents Fonction : Certains insectes injectent des miARN capables de réguler négativement l’expression de gènes de la plante, en ciblant des ARN messagers spécifiques. Exemple : Des études sur les pucerons ont montré que leurs miARN peuvent réprimer des gènes impliqués dans la défense ou la morphogenèse foliaire c. Phytohormones et leurs analogues provenant de la plante hôte mais détournées dans leur fonction Auxines : Stimulent la division cellulaire et l’élongation, souvent responsables de la croissance initiale de la galle. Cytokinines : Favorisent la différenciation cellulaire et la formation de tissus spécialisés (comme les poils ou les vaisseaux conducteurs dans la galle). Gibbérellines : Peuvent être impliquées dans l’allongement des cellules. Acide abscissique (ABA) : Parfois détourné pour modifier la réponse au stress ou la fermeture des stomates. 2. Réponse de la plante : reprogrammation du développement Les signaux injectés par l’insecte déclenchent une cascade de réactions dans la plante, aboutissant à la formation de la galle. Voici les étapes clés : .a Activation de voies de signalisation Voie des MAP kinases : Impliquée dans la transduction du signal de stress et la division cellulaire. Voie de l’éthylène : Souvent réprimée pour éviter la sénescence ou la nécrose des tissus. Voie du calcium : Les flux de Ca²⁺ intracellulaires peuvent activer des facteurs de transcription liés à la croissance. b. Modification de l’expression génique Gènes de développement : Des gènes normalement actifs dans les méristèmes (zones de croissance) ou les organes floraux sont réactivés dans les cellules de la galle. Par exemple : Gènes KNOX (impliqués dans le maintien des cellules souches végétales). Gènes WUSCHEL (régulateurs de la différenciation). Gènes métaboliques : Augmentation de l’expression de gènes codant pour des enzymes de biosynthèse de nutriments (sucres, acides aminés), qui nourriront l’insecte. c. Remodelage du cytosquelette et de la paroi cellulaire Protéines du cytosquelette (actine, tubuline) : Leur organisation est modifiée pour permettre une division cellulaire asymétrique ou une expansion directionnelle. Enzymes de modification de la paroi (expansines, xyloglucane transférases) : Permettent l’expansion et la différenciation des cellules. 3. Spécificité de la galle : un dialogue évolutif La forme et la structure de la galle dépendent d’une co-évolution entre l’insecte et la plante : L’insecte a développé des effecteurs capables de cibler des gènes clés de la plante hôte. La plante peut, en retour, évoluer pour résister à ces manipulations (par exemple, en modifiant ses récepteurs hormonaux ou en activant des défenses ciblées). 4. Outils pour étudier ces mécanismes Les chercheurs utilisent : Le séquençage ARN (RNA-seq) : Pour comparer l’expression des gènes dans les tissus sains et galligènes. La CRISPR-Cas9 : Pour inactiver des gènes candidats et observer l’effet sur la formation de la galle. La microscopie confocale : Pour visualiser les modifications du cytosquelette ou la localisation des hormones En résumé La galle est bien le résultat d’une manipulation génétique fine par l’insecte, qui exploite les voies de développement de la plante pour créer une structure adaptée à ses besoins. Besoin de protection Besoin de nutriment ‘les galles deviennent des greniers d’ amidon Ce n’est pas une croissance anarchique, mais une réorganisation contrôlée du tissu végétal, presque comme un "organe" supplémentaire imposé par l’insecte. Références: Guides des Galles de France et d'Europe Le texte est issu d'une discussion avec Chat Mistral IA. Dominique

-
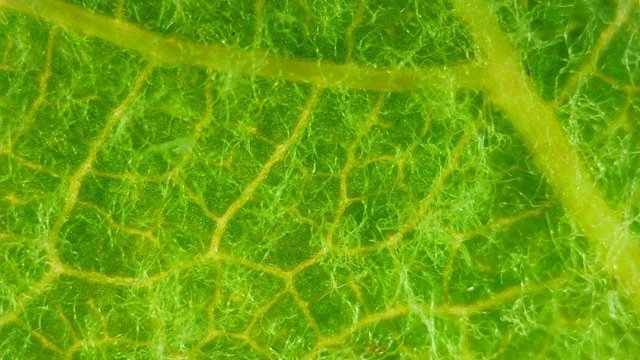
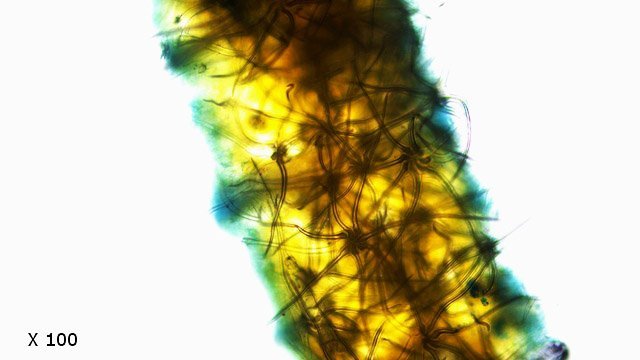
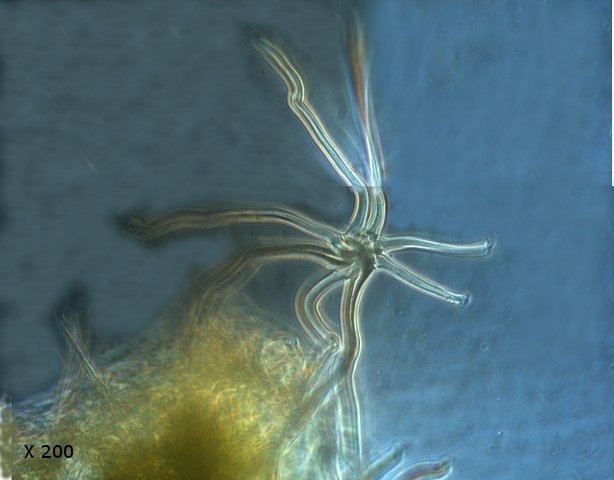
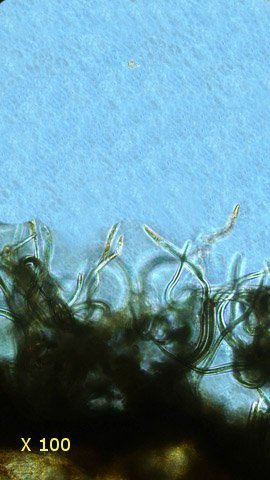
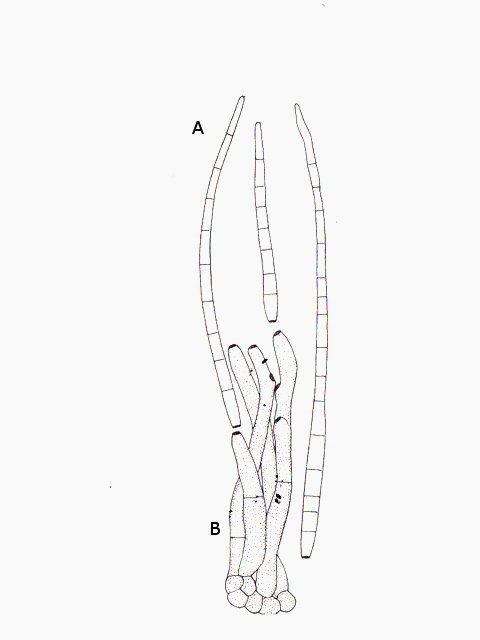
Tilleul fungus automne A l’automne les feuilles se couvrent de taches de couleurs variées , souvent brunes à noires .Une partie de ces taches correspond à la mort programmée des cellules constituant la feuille conséquence de l’abscission , une partie provient de l’ installation de moisissures ou comme on l’a vu de galle Le tilleul perd ses feuilles ce qui nous permet de les découvrir aisément Feuille de tilleul : Une coupe de la feuille est réalisée : Vue de dessus , ce qui sur la photo macro correspond au chevelu bien visible : Et en contraste de phase : Deux types d’ hyphes peuvent être isolés: Des hyphes fertiles surmontées par une ou plusieurs spores De hyphes non conidiogénes.;Ces hyphes semblent se rassembler en bouquets de 3 à 8 hyphes et sont responsables de l’ aspect feutré de la colonie vue en épiscopie. A partir de ces images il est possible de réaliser le schéma suivant : A Conidiophore B Spores C Hyphes réunies en faisceaux D Poils stériles Comme d’ habitude je ne suis pas capable de donner un nom à cette moisissure mais j’ en retire cependant le grand plaisir de découvrir une forme inattendue. Dominique.
-
Merci beaucoup de ton aide Jmaffert Mais c'est un sujet que je ne possède pas bien mais j' aurai dû me méfier en regardant la coupe de cette gale ( la zone creuse centrale ne pouvait pas être compatible avec un fungus), je n ai pas utiliser la recherche du diagnostic différentiel en constatant certaines incohérences . Sur wikipedia on nous enseigne que "Neuroterus quercusbaccarum, parfois appelé cynips galle-groseille ou cynips galle-lentille[1], est une espèce de la famille des « guêpes à galles » ou « mouches à galles », les Cynipidae. Cet insecte est responsable de la formation de galles sur les chênes à feuilles caduques : des galles sphériques à l'allure de groseilles sur les inflorescences mâles ou des galles à l'allure de lentilles sous la face inférieure des feuilles." Merci Tryphon pour l' article que tu nous as indiqué - cet article est passionnant est révèle la complexité du monde vivant . Amicalement
-
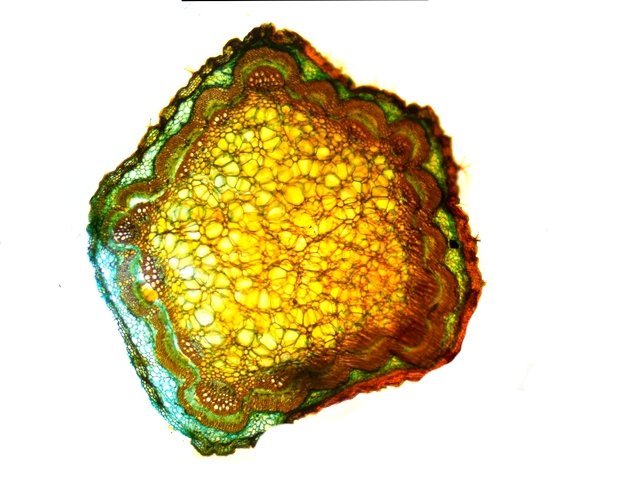
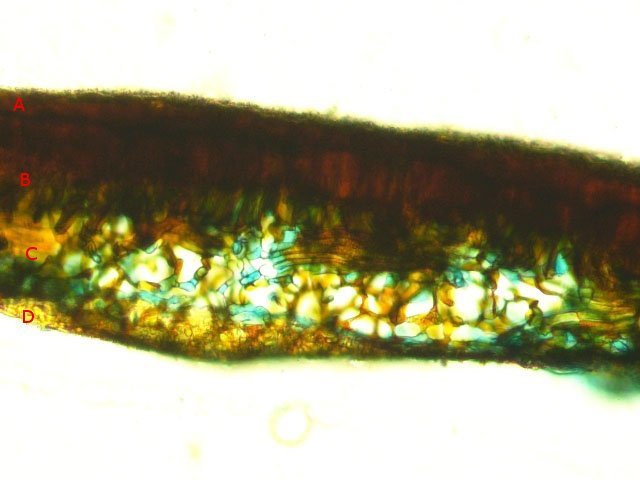
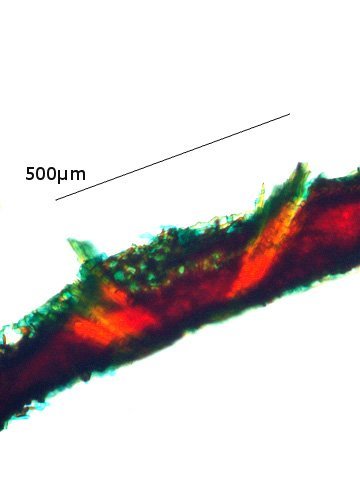
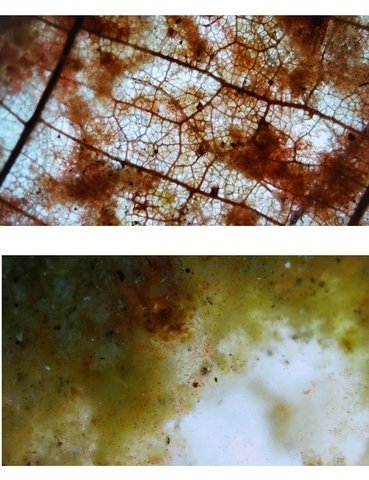
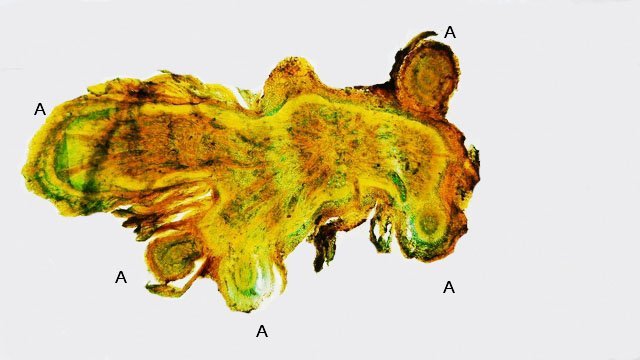
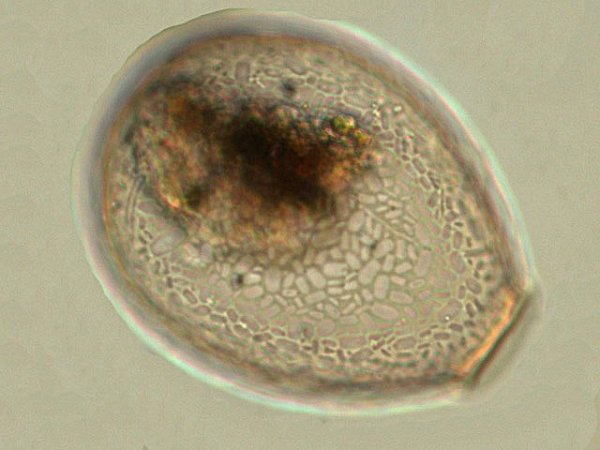
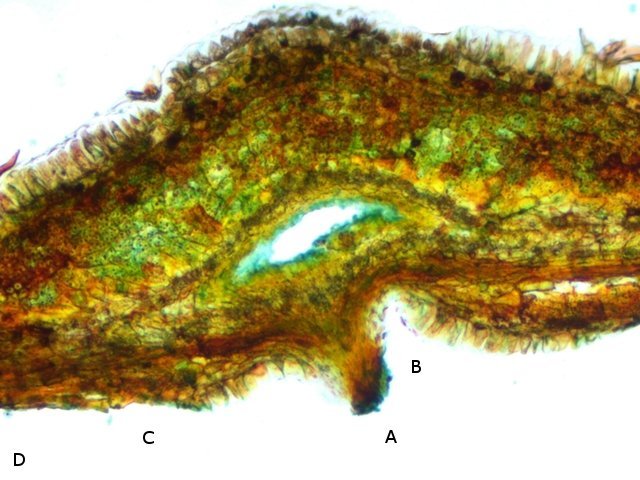
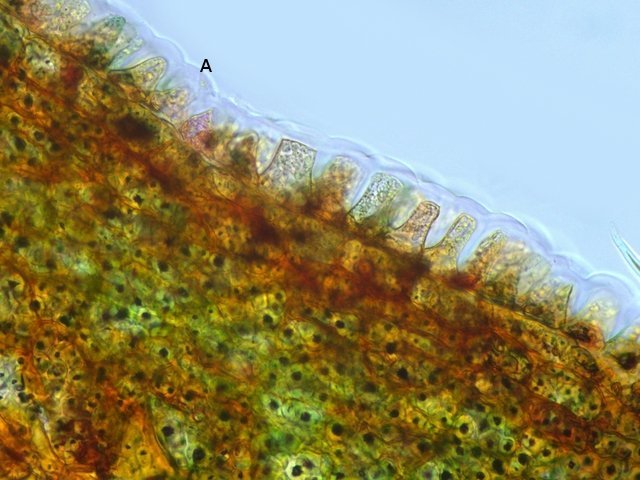
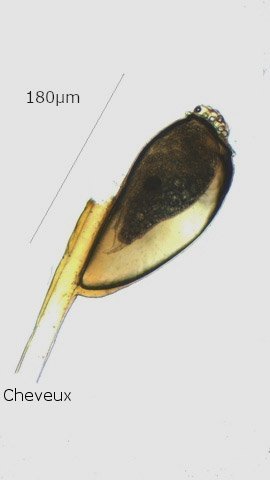
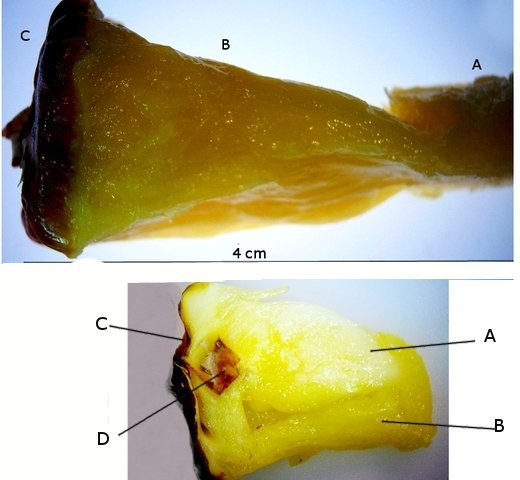
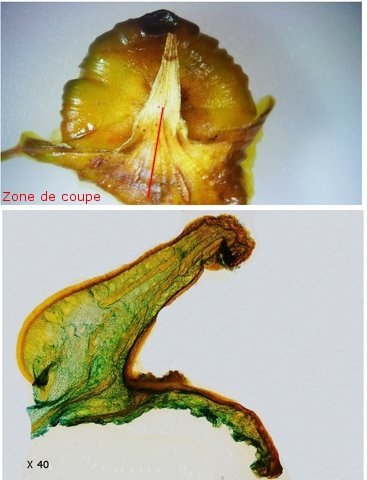
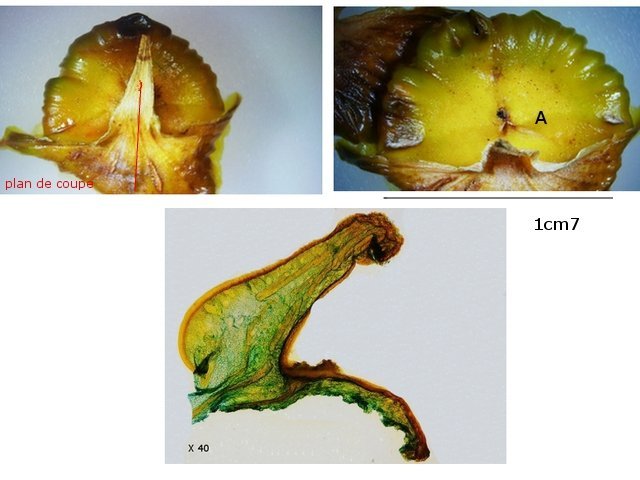
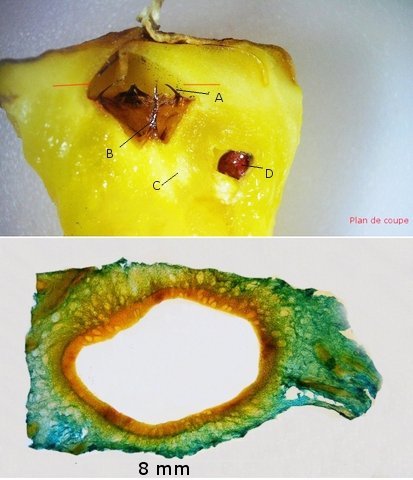
Discomycètes sur feuille de chêne ? Note de correction — Les structures initialement décrites comme Discomycètes se sont révélées être des galles de cynips (probablement Neuroterus quercusbaccarum). L’hypothèse fongique a donc été abandonnée, mais les observations et coupes restent utiles à titre documentaire. Tous les ans en automne les feuilles du vieux chêne au bas de la route se couvrent de taches - Toutes n’ont par la même forme indice ni la même couleur indices d'attaques multiples réalisées par des variétés de guêpes différentes. 3 formes sont présentes: . Chaque petit rond fait autour de 4 mm de diamètre et il est très facile de les décoller du limbe de la feuille. Sur cette première photo les formations sont marron-clair et aplaties. Il existe une autre forme dont les rebords sont bien marqués. Et une troisième forme plus claire et plus bombée Regard sur la face inférieure: En A est la seule zone qui a un contact avec le limbe de la feuille. Une coupe est réalisée : A Fixation sur le limbe de la feuille – B Limbe de la feuille C Coupe du parenchyme - une cavité est bien délimitée dans la base de la galle mais cette cavité est vide . D Ornementations Reste à trouver le nom de cette formation ou le nom de ces formations puisqu' il en a été trouvé 3 . Dominique
-
Cercosporiose Cercospora beticola Pas loin de la maison il y a un immense champ de Betteraves fourragères. Les betteraves sont énormes - En faisant bien attention on constate que les feuilles des betterave du champ sont parfaites homogènes - volumineuses .Par contre sur la bande de limite du champs et sur une largeur d’un mètre les feuilles sont petites tachées parfois sèche et marron La séparation entre les deux zones est bien rectiligne : le pulvérisateur agricole a donc laissé 1 mètre sans traitement Etat des feuilles atteintes : Après grossissement des taches : Coupe de la partie atteinte : La zone de transition La structure normale de la feuille a été totalement détruite . Comparaison avec la partie saine de la feuille : Diagnostic : Atteinte des feuilles de betteraves par un fungus Cercospora beticola . A – Conidies ( produites ici en chaînette ) B -- Conidophores En lumière polarisée il est constaté une importante charge minérale des feuilles de betterave . Pour en savoir plus : La cercosporiose est une maladie fongique causée par des champignons du genre Cercospora. Elle touche principalement les plantes, en particulier les cultures agricoles comme la betterave sucrière, la carotte, la tomate, le soja, le riz, le maïs, et certaines plantes ornementales. Symptômes principaux Taches foliaires : Petites taches circulaires, souvent grises ou brunes, entourées d’un halo jaune. Nécrose : Les taches s’élargissent et finissent par sécher, provoquant la chute des feuilles. Affaiblissement de la plante : Réduction de la photosynthèse, baisse des rendements, voire mort de la plante en cas d’infection sévère. Conditions favorables Humidité élevée (rosée, pluie, irrigation) Températures modérées (20–30 °C) Mauvaises pratiques culturales (rotation insuffisante, résidus infectés) Exemples de cultures touchées Betterave : Cercosporiose de la betterave (Cercospora beticola) Tomate : Cercosporiose de la tomate (Cercospora fuligenoides) Soja : Cercosporiose du soja (Cercospora sojina Classification Catalogue of Life Règne Fungi Embranchement Ascomycota Classe Dothideomycetes Ordre Capnodiales Famille Mycosphaerellaceae Genre Cercospora Espèce Cercospora beticola 1. Traitements fongicides chimiques Les fongicides sont souvent utilisés en prévention ou dès l’apparition des premiers symptômes. Trois spécialités présentent de bonnes performances Produits à base de cuivre Prothioconazole+fluopyramil Difénoconazole + fenpropidine Les seuils de traitements sont atteints lorsque 1% ( 5 % en bord de mer )des feuilles présentent des symptômes 3. Méthodes culturales et préventives Rotation des cultures : Ne pas replanter la même espèce sur une parcelle infectée pendant 3–4 ans. Choix de variétés résistantes : Certaines variétés de betterave, soja ou tomate sont moins sensibles. Gestion de l’irrigation : Éviter l’arrosage du feuillage, privilégier le goutte-à-goutte. Désherbage : Limiter les adventices qui peuvent héberger le champignon. Élimination des résidus infectés : Brûler ou enfouir profondément les débris végétaux après récolte. NB Il y a peu de différences entre variétés fourragères et variétés potagères pour les betteraves , aussi autre fois ( mais peut être dans le future )en cas de pénurie, on utilisait les variétés fourragères plus productives pour la consommation humaine. Cela a été le cas pendant les deux dernières guerres mondiales en Europe.en dans les périodes de famine en Irlande. Ref La France agricole N° 4120 Dominique .
-
Bonjour Tryphon Ton histoire fait revenir dans ma mémoire des souvenirs d’enfance où les poux étaient appelés des Totos . Cette expression a disparu de nos jours mais le fait que tu réveilles cette expression de jeunesse est tout à fait agréable. Amicalement Dominique
-
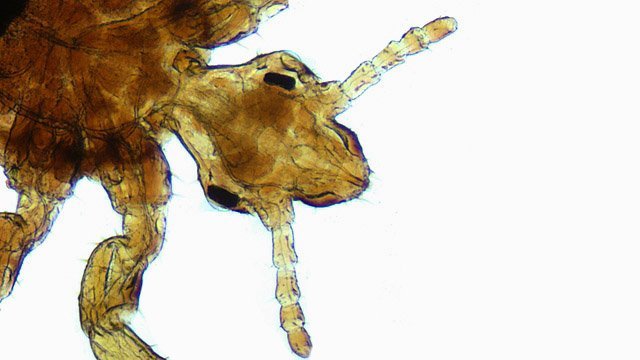
Poux Pediculus humanus capitis L’ année scolaire a commencé il y a 5 jours et sur les cheveux de ma petite fille on découvre des poux Un prélèvement est réalisé : Sur cette photo le repas sanguin est bien visible ( lors du prélèvement sur la tête il était rouge ) A noter la quantité d’ œufs contenus dans le sac à œufs . Un pou de tête femelle peut pondre entre 6 et 10 œufs par jour. Sa durée de vie est d'environ 30 jours .Pendant sa vie, une femelle pou peut donc produire environ 200 lentes. : Le pou a un système piqueur suceur rétractable » le stylet » ( ce qui est le cas de la photo )la salive contient un anticoagulant. Les démangeaisons sont causées par une réaction allergique à la salive injectée par le pou lors de la morsure. Les zones les plus fréquemment affectées incluent la nuque, derrière les oreilles et la base des cheveux. Les piqûres peuvent également entraîner des réactions cutanées inflammatoires, des éruptions cutanées et des petites bosses rouges sur la peau. En cas de grattage intense, des infections bactériennes secondaires peuvent se développer, provoquant des lésions cutanées et des plaques de peau irritée. Le pou se déplace très vite il court sur la peau à la vitesse de 23 cm à la minute par contre il ne saute pas comme la puce. Les lentes : Temps d'incubation des lentes Éclosion : Les lentes éclosent généralement 7 à 10 jours après avoir été pondues par le pou femelle. Durée de vie : Une fois éclose, la larve (appelée nymphe) met environ 7 à 10 jours supplémentaires pour devenir un pou adulte capable de se reproduire. Les lentes vides (après éclosion) restent souvent accrochées aux cheveux et peuvent être confondues avec des pellicules, mais elles sont plus difficiles à enlever. Les poux ne survivent pas plus de 24 à 48 heures hors du cuir chevelu. **************************** 1. Classification et types Le pou est un insecte ectoparasite (vivant à l'extérieur du corps de son hôte). Chez l'être humain, on distingue trois espèces principales : Les poux sont des parasites exclusifs de l'homme Le pou de tête (Pediculus humanus capitis), le plus fréquent, qui vit sur le cuir chevelu. Le pou de corps (Pediculus humanus humanus), qui vit sur les vêtements et se nourrit sur la peau. Le pou de pubis (Phtirus pubis), plus connu sous le nom de morpion, qui vit dans les poils pubiens. 2. Le pou de tête : cycle de vie et caractéristiques Le pou de tête a un cycle de vie en trois stades : la lente, la nymphe et le pou adulte. La lente (l'œuf) Les femelles pondent 4 à 10 œufs par jour. Les lentes sont de minuscules œufs ovales, de couleur grisâtre ou beige, et sont fermement fixées à la base du cheveu, à moins d'un centimètre du cuir chevelu, grâce à une sorte de colle (le cément). Elles éclosent au bout de 7 à 10 jours. Les lentes vides, plus claires, peuvent rester accrochées au cheveu pendant des mois. La nymphe (le jeune pou) C'est le stade qui suit l'éclosion de la lente. La nymphe ressemble au pou adulte, mais en plus petit. Elle subit trois mues successives avant de devenir un pou adulte. Ce stade dure environ 10 à 15 jours. Le pou adulte Il mesure de 2 à 4 mm de long. Il a six pattes, chacune munie de pinces qui lui permettent de s'accrocher fermement aux cheveux. Le pou est un insecte hématophage, ce qui signifie qu'il se nourrit exclusivement de sang humain, qu'il prélève plusieurs fois par jour sur le cuir chevelu. Il ne peut pas survivre plus de 2 jours en dehors de son hôte. La femelle pou peut vivre de 1 à 2 mois. 3. Transmission et symptômes La transmission se fait principalement par contact direct de tête à tête. La transmission indirecte via des objets (bonnets, peignes, brosses, écharpes) est possible, mais moins fréquente car les poux ne survivent pas longtemps hors de l'hôte. Les poux se déplacent en rampant. Le symptôme le plus courant est une forte démangeaison du cuir chevelu (prurit), causée par la salive du pou. 4. Résistance Les poux sont assez résistants. Ils peuvent survivre sous l'eau en fermant leurs orifices respiratoires. Ils peuvent également survivre à des températures élévées tant qu'ils sont en contact avec la peau de leur hôte, qui agit comme un thermorégulateur. De plus, certaines populations de poux ont développé une résistance aux insecticides (pédiculicides) couramment utilisés 1. Classification taxonomique du pou Classification du pou de tête Embranchement Arthropoda Classe Insecta Ordre Phthiraptera Sous-ordre Anoplura Famille Pediculidae Genre Pediculus Espèce Pediculus humanus capitis (Pou de tête) Il existe un grand nombre de produits anti poux dont l’ efficacité est parfois discutable - Un jour je vois une famille dont tous les membres sont atteints d’ une gale Le responsable de la gale humaine est un acarien microscopique appelé Sarcoptes scabiei var. hominis (ou simplement "sarcopte"). Ce parasite est spécifique à l’humain et ne survit que quelques jours en dehors de la peau.On traite cette maladie par des comprimés d' Ivermectine . Je revois la famille quelques jours plus tard pour vérifier l’ efficacité du traitement et ils me disent « Docteur on a essayé tous les traitements possibles contre les poux et depuis la prise de votre médicament nous n’ avons plus rien ni gale ni poux ". Depuis pour le infestations importantes par les poux je le prescris avec efficacité. Dominique
-
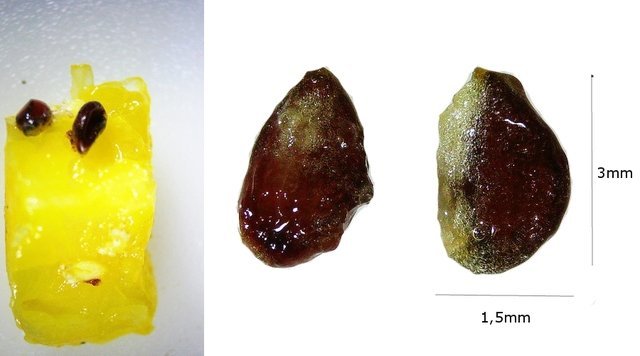
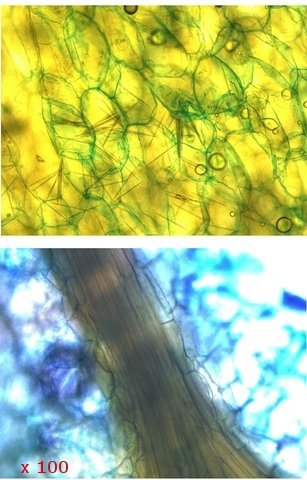
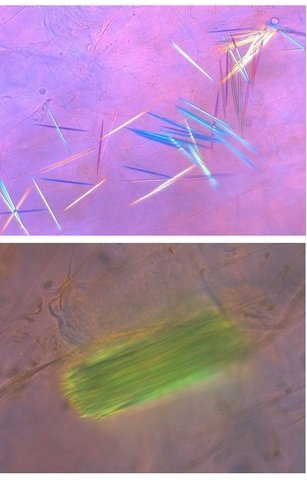
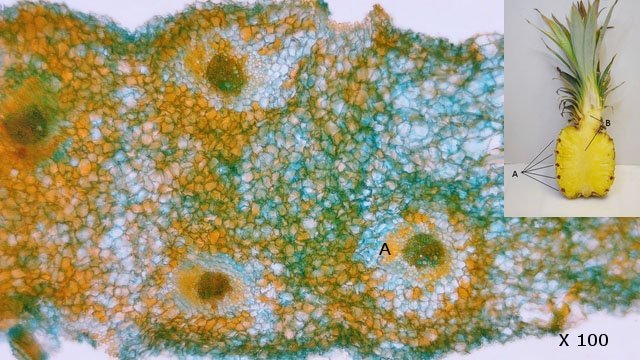
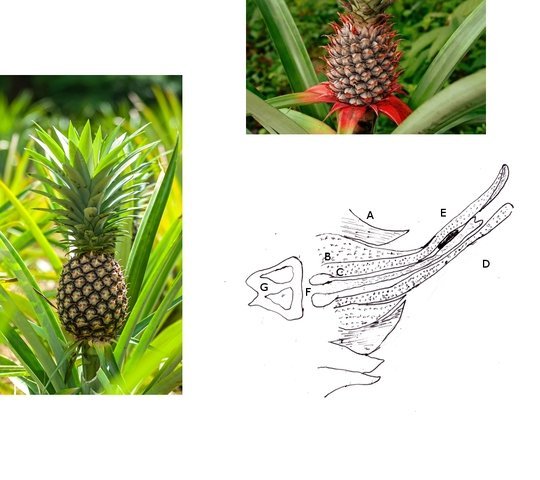
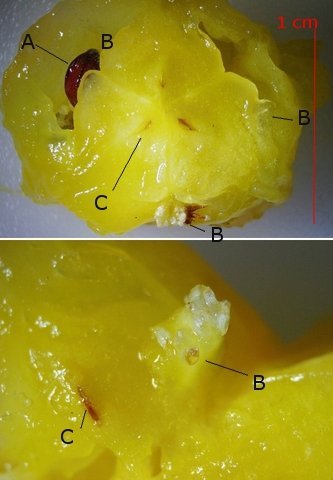
Ananas structures histologiques Lorsque l’ on regarde un ananas en Normandie on se place devant la fin d’ un processus celui de la fructification . La plante est une monocotylédone herbacée et pérenne et hermaphrodite. Sa caractéristique est le développement d’une tige entourée de feuilles qui va porter une inflorescence surmontée d’un développement de feuilles linéaires . A Bractée B Sépale C Pétale D Pistil E Etamine F Calice G ovaire Une fois cette inflorescence devenue fruit la plante va recommencer à partir d’ un bourgeon axillaire de la tige initiale le même processus. Cette nouvelle tige adventive portera un nouveau fruit souvent plus petit -( de ce fait en culture industrielle on limite la croissance de la plante à un parfois deux fruits). **Sur la photo du haut on constate que la production se fait successivement à partir de la tige centrale et de branches adventives (ici future 3 éme récolte **A calices de quelques fleurs B axe central. Ce Pseudo fruit est donc obtenu par la coalescence de chacun des petits fruits qui se sont développés dans l inflorescence Cette Inflorescence est faite de fleurs sessiles (sans pédicelle) sur un axe unique. La floraison commence à la base et se poursuit vers l'apex. La fleur évolue : Les pétales vont disparaitre les sépales et les bractées vont se souder pour fermer la partie fructifère de la fleur Cette partie fructifère est le gynécée de la fleur . Photo du haut A Tige centrale B Zone du Mésocarpe C couverture par repliement des bractées formant l’Epicarpe. .La coupe est faite au milieu du fruit . Photo du bas A gynécée (paroi externe de l’ovaire + ovaire + ovule ) . B endocarpe .C bractées de couverture . D calice résiduel . La coupe met en évidence une forte lignification de la zone du liber.des bractées repliées.renforçant la protection du fruit sous-jacent. A noter sur la photo suivante la présence d’un orifice qui sert à émettre un abondant nectar provenant des glandes nectarifères situées dans l’ovaire. Sous ce couvercle protecteur formé par le repliement des bractées charnues se trouve le calice de la fleur. ***On retrouve en A des restes secs des étamines B les restes secs du pistil C la zone de l’endocarpe qui entoure l’ovaire D le développement d’une graine sur la zone ovulaire. Sur la photo du bas il est mis en évidence une forte lignification de la paroi interne du calice. . L’ovaire autour de 2 cm : Photo du haut :Sur cette photo l’ovaire est formé par 3 loges carpellaires contenant un placenta ici . il n’ y a qu’un seul placenta qui s’est développé En C canal nectarifère dont l’ origine est située sous les ovaires Sur la photo du bas Le placenta développé ( B ) est surmonté de plusieurs ovules qui donneront naissance aux graines On en compte 5 sur cette photo. Chaque Gynécée est entouré d’une importante masse de tissu parenchymateux qui a un rôle conjonctif entre les différentes fleurs constitutives de l’ananas Dans le cas d’ ananas il n’ y a pas de Péricarpe individualisable. C’est donc une infructescence ( c’est-à-dire la réunion dans une seule masse tissulaire de nombreux petits fruits individuels ) - ce processus est connu chez la fraise . Évolution des ovules Une très petite partie seulement des ovules va évoluer vers la formation de petites graines L’ ovaire a une position infère c’est-à-dire que l’ovaire est situé au-dessous du niveau d’insertion ,sur le réceptacle , des autres pièces florales ( sépales -pétales ) on dit que la fleur est épigyne ). Les cultivateurs d'ananas, pour empêcher la fécondation des fleurs, n'ont en fait pas grand-chose à faire : ils n'ont pas besoin de s'arranger pour l'éviter car la variété d'ananas la plus couramment cultivée, l'ananas Cayenne Lisse, est parthénocarpique.. Cela signifie que le fruit se développe sans avoir besoin de fécondation Mais il y a surtout de nombreuses variétés qui sont cultivées dans des zones présentant peu de colibris et de chauves-souris Il y a donc peu de graines L’ananas tient sa réputation de l’important développement de son parenchyme intermédiaire le Mésocarpe Le mésocarpe constitue la partie intermédiaire du fruit communément appelée pulpe quand il s'agit de fruits charnus. Il dérive de la transformation du parenchyme de la paroi de l'ovaire.Aspect microscopique: La pulpe est faite d’un tissu constitué de grandes cellules, lui-même traversé par les cellules fibreuses assurant sa résistance L’ananas est réputé pour ses raphides l’image ci-dessus donne une idée de l’importance de ces aiguilles cristallines – les raphides font autour de 20 µm Il en existe deux aspects: Soit sous forme d’aiguilles isolées soit sous forme de petits fagots. Il existe sur MikrOscOpia plusieurs présentations: https://forum.MikrOscOpia.com/topic/20068-l-ananas-un-fruit-qui-ne-manque-pas-de-piquant/#comment-83216 les raphides de l’ananas . https://forum.MikrOscOpia.com/topic/1527-ananas-comosus-ananas/#comment-6282 la feuille. Au centre de l’ananas ( B ) se trouve un axe rigide qui assure le maintien rigide de toute la plante . Cet axe central est dur et très rigide il fait un diamètre de 2 cm - il est la colonne vertébrale de la plante sur laquelle vont se fixer les multiples petites fleurs et les feuilles .En A se positionnent les nombreux axes vasculaires. Sa lignification est importante;De cette tige naissent aussi de nombreuses plantules qui repiquées vont permettre la multiplication végétative de l 'ananas. La reproduction des pieds d’ananas Les ananas se reproduisent principalement par division des rejets à la base de la plante ou par bouturage de la couronne du fruit, moins fréquemment par graines, qui sont lentes à germer. La méthode la plus courante consiste à couper la couronne, la tremper dans l'eau, puis la replanter dans une terre bien drainée Bien que les plants d’ananas soient des plantes vivaces, ils vivent rarement plus de six ans s’ils produisent des fruits. Après la production de fruits, la qualité de la plante diminue. Cependant, la plante commence à produire des pousses qui peuvent être multipliées pour former de nouvelles plantes. Pourquoi les ananas donnent souvent une sensation de picotement dans la bouche ? La sensation de picotement ou de brûlure que l'on ressent en mangeant de l'ananas est bien due à la bromélaïne. La bromélaïne est un complexe d'enzymes protéolytiques, ce qui signifie qu'elle a la capacité de décomposer les protéines. Lorsque vous mangez de l'ananas, cette enzyme entre en contact avec les tissus de votre bouche, de votre langue et de vos lèvres, qui sont composés de protéines. La bromélaïne commence alors à "digérer" ces protéines, provoquant cette sensation de picotement désagréable. La concentration de bromélaïne est plus élevée dans la tige et le cœur de l'ananas, ainsi que dans les fruits moins mûrs. C'est pourquoi un ananas bien mûr et consommé loin du cœur piquera généralement moins. En plus de cette particularité, la bromélaïne a également de nombreuses propriétés bénéfiques pour la santé, et elle est d'ailleurs utilisée dans l'industrie pharmaceutique et dans certains compléments alimentaires pour ses vertus anti-inflammatoires et digestives. Classification Taxonomique/ Règne : Plantae (Plantes) Division : Tracheophyta (Plantes vasculaires) Classe : Liliopsida (Monocotylédones) Ordre : Poales Famille : Bromeliaceae (Broméliacées) Genre : Ananas Espèce : Ananas comosus Référence The pineapple :botany ,production as uses par Kenneth G Zimmer et Daniel P Bartholomew Microscope utilisé le Tomlov microscope à écran L’objectif est un zoom ce qui rend difficile la précision de l’agrandissement Dominique
-
Méristème Ronce croissance du turion.
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
Bonsoir Tryphon Merci pour la vidéo bien que je doute de l' action sur le pied mère de la ronce -- je n'ai aucune expérience de la technique - Pour l' instant le sécateur reste une technique qui maîtrise bien la prolifération des ronces sans bien sûr en venir à bout . Pour le grossissement et les précisions techniques de la prise des photos je reconnais ma grande négligence - j' aime surtout raconté l'histoire ; je vais faire un effort pour le prochain article qui sera encore de la botanique . Amicalement Dominique -
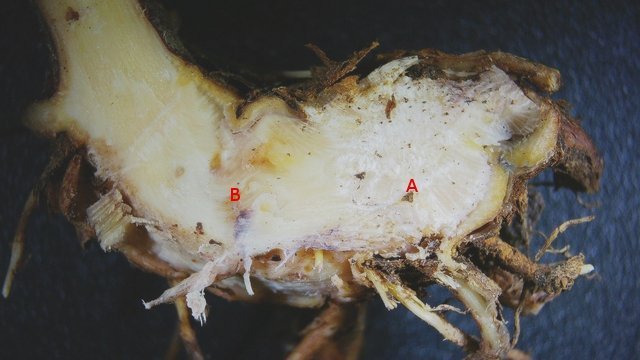
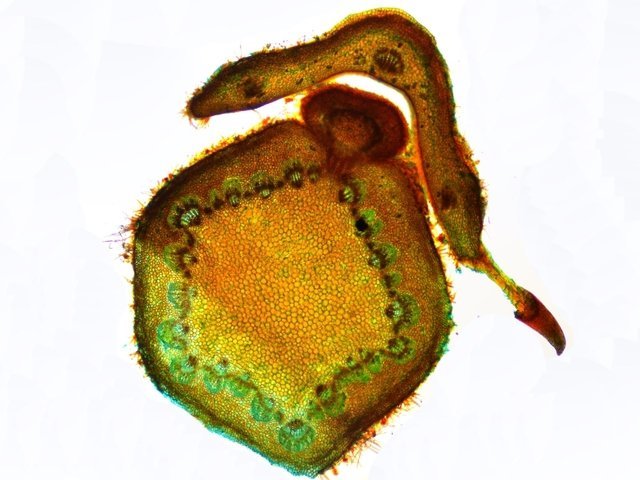
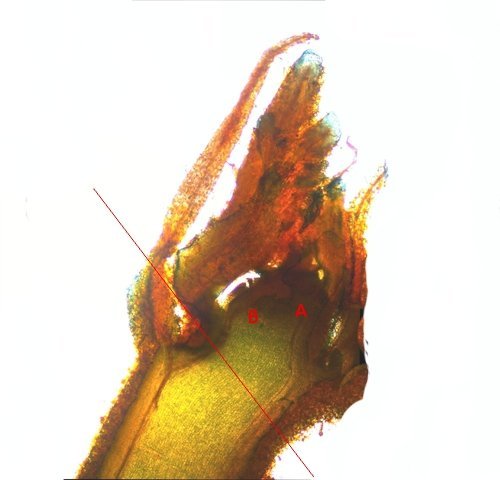
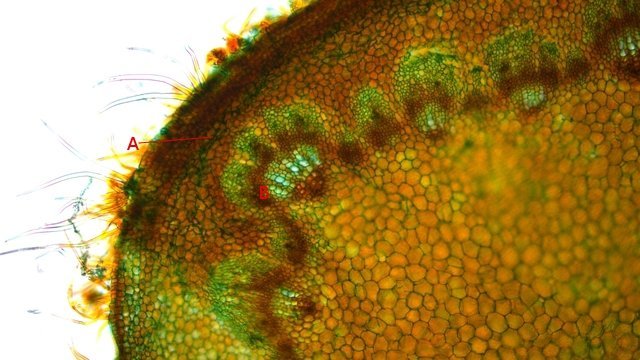
Méristème Ronce croissance du turion. Dans le jardin j’ai beaucoup de tiges de ronce - que je coupe tous les ans à la base ce qui permet d’assister à la repousse d’une nouvelle tige l’ année suivante. Chez la ronce (famille des Rosacées), le turion est une tige feuillée dépourvue de fleur, longue et vigoureuse, émise par la souche, et se ramifiant l'année suivante en produisant des rameaux florifères. Cette tige ligneuse bisannuelle peut être dressée ou retombante . (Le turion est donc un bourgeon enterré encore appelé œil ; il donne naissance chez certaines plantes vivaces comme les asperges à une jeune pousse, qui apparaît à la période de végétation sur les racines, rhizomes ou tubercules). On constate que les turions développent d’ abord des feuilles à leur extrémité distale. A – Rhizome B – Bourgeon du turion qui s’est développé à partir du rhizome Ce qui est remarquable chez les ronces c’est la vitesse de croissance de cette tige de l’année - qui peut atteindre plus de 30 cm par semaine. Il était intéressant de regarder l’extrémité distale de cette tige là où se situe le méristème apicale responsable de la croissance en longueur. Définition : Les méristèmes sont une catégorie générale de tissus végétaux. Ce sont des zones de cellules indifférenciées qui se divisent activement par mitose. On peut les considérer comme les "cellules souches" de la plante. Leur rôle principal est de permettre la croissance de la plante. Il faut bien garder conscience que les cellules du méristème ne constituent pas de grandes plages cellulaires mais seulement quelques lignes plus ou moins étendues qui se confondent dans la masse des cellules filles différentiées technique Coloration des coupes Acridine 1minute /Bleu Alcian 1 minute. L extrémité distale : A zone du méristème apical . B zone des méristèmes secondaires. C zone de la coupe - toutes les structures sont déjà en place. – on se situe à ( 5 mm de la partie distale ) . On constate donc que les méristèmes les plus distaux sont des méristèmes caulinaires : Méristème Apical Caulinaire (MAC) : Localisation : Au sommet de la tige, dans le bourgeon terminal ou les bourgeons axillaires. Rôle : Il est à l'origine de la tige, des feuilles et des fleurs. Il contient un groupe de cellules souches qui se divisent pour produire de nouvelles cellules, qui se différencient ensuite en différents tissus primaires de la tige (épiderme, cortex, tissus vasculaires primaires). Donc la plante en même temps qu’elle entame sa croissance en longueur développe son enveloppe de protection (primordiums foliaires) qui deviendra des feuilles. Méristèmes secondaires : A ce niveau les méristèmes vont en très peu de temps se différentieren méristèmes secondaires Méristèmes latéraux Ces méristèmes sont responsables de la croissance en épaisseur (croissance secondaire) des plantes, principalement chez les gymnospermes et de nombreuses dicotylédones. Ils sont disposés cylindriquement le long de la tige et de la racine. On trouve Méristème latéral ou Cambium Subéreux (ou Phellogène) : Localisation : Généralement situé sous l'épiderme ou dans le cortex. Rôle : Il produit le suber (liège) vers l'extérieur et le phelloderme (un tissu parenchymateux) vers l'intérieur. L'ensemble suber, phellogène et phelloderme forme le épiderme, qui remplace l'épiderme comme tissu protecteur chez les plantes à croissance secondaire. Méristème vasculaire ou Cambium Vasculaire : Localisation : Entre le xylème et le phloème. Rôle : Il produit du xylème secondaire (bois) vers l'intérieur de la plante et du phloème secondaire (liber) vers l'extérieur. C'est l'activité du cambium vasculaire qui est responsable de l'augmentation du diamètre des troncs d'arbres et de la formation des cernes de croissance annuels. A zone des Cambium subéreux (méristème subéreux) . B zone des Cambium vasculaire (méristème vasculaire). Aspect des cellules du méristème Pour les méristèmes apicaux (situés aux extrémités des tiges et des racines), on observe souvent une organisation en couches ou en zones distinctes, comme le centre quiescent (une zone de cellules qui se divisent peu, servant de réservoir de cellules souches) entouré de cellules qui se divisent activement. Cette organisation est loin d'être une "large plage" uniforme. Mais plutôt quelques couches de cellules aux caractéristiques particulières Caractéristiques histologiques des méristèmes : Coloration vert méthyle .. Coloration Safranine pour la mise en évidence des noyaux: Description des cellules des méristèmes: Petites cellules, isodiamétriques et étroitement serrées : Les cellules du méristème sont généralement de petite taille, de forme arrondie ou polyédrique, et ne présentent pas d'espaces intercellulaires. Elles sont donc très compactes. Parois cellulaires minces : Les parois de ces cellules sont fines et composées principalement de pectine et de cellulose, ce qui facilite leur division et leur croissance. Noyau proéminent : Chaque cellule a un grand noyau qui occupe une part importante de son volume, ce qui est un signe d'une intense activité métabolique et de division cellulaire. Le rapport nucléocytoplasmique (rapport entre le volume du noyau et celui du cytoplasme) est élevé. Cytoplasme dense : Le cytoplasme est abondant et dense, riche en ribosomes et en mitochondries, ce qui témoigne d'une forte synthèse de protéines et d'une grande activité métabolique. Vacuoles réduites ou absentes : Contrairement aux cellules matures, les cellules du méristème ont de très petites vacuoles ou n'en ont pas du tout. La majeure partie de leur volume est consacrée au noyau et au cytoplasme, essentiels à la division. Absence de substances de réserve : Les cellules du méristème n'ont pas de substances de réserve comme l'amidon. Elles utilisent l'énergie et les nutriments pour la division et non pour le stockage. A noter que : Pour les méristèmes apicaux (situés aux extrémités des tiges et des racines), on observe souvent une organisation en couches ou en zones distinctes, comme le centre quiescent (une zone de cellules qui se divisent peu, servant de réservoir de cellules souches) entouré de cellules qui se divisent activement. Cette organisation est loin d'être une "large plage" uniforme. En conclusion, si on observe une coupe histologique d'une plante, les méristèmes ne se présentent pas comme de vastes étendues de cellules, mais plutôt comme des zones localisées de cellules aux caractéristiques embryonnaires, compactes et en divisions actives, clairement distinctes des tissus matures environnants. En quelques jours la tige a pris son aspect histologique caractéristique de la tige de ronce . ,Coupe axiale et Coupe longitudinale d'une tige à 1 cm de la zone apicale: Toutes ces modifications histologiques se font à grande vitesse c’est-à-dire en quelques heures d’où la croissance rapide des turions. Une plante ne semble pas bouger à notre niveau d’observateur humain mais les processus en cours sont déroutants . NB Chez les poacées ( herbe ) il existe des Méristèmes Intercalaires. Bien que moins mis en avant que les apicaux et secondaires, les méristèmes intercalaires sont également importants, particulièrement chez les graminées. Localisation : À la base des entrenœuds ou des feuilles. Rôle : Ils permettent une croissance rapide en longueur à partir de ces points, ce qui explique pourquoi les pelouses peuvent repousser rapidement après avoir été tondues. Référence: NB aide pour le texte en partie avec Gemini IA Dominique .
-
-
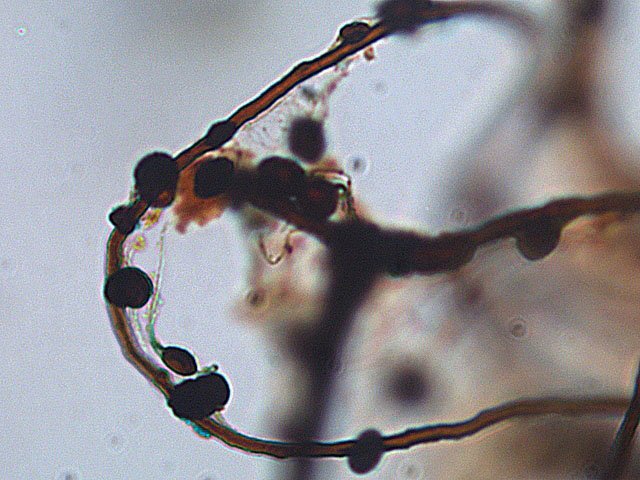
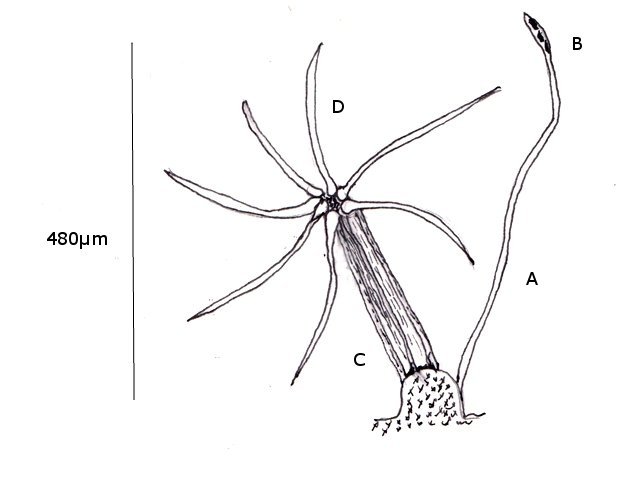
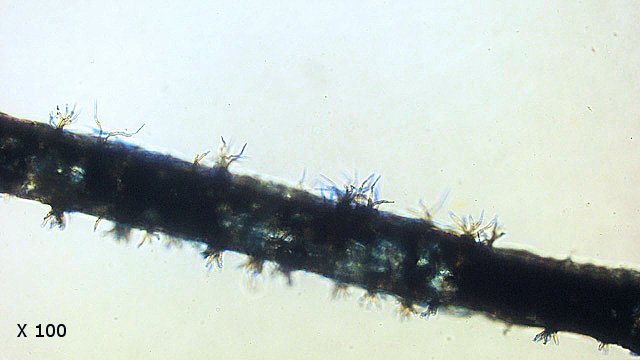
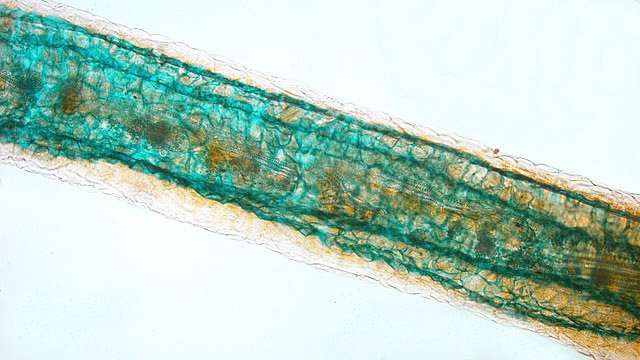
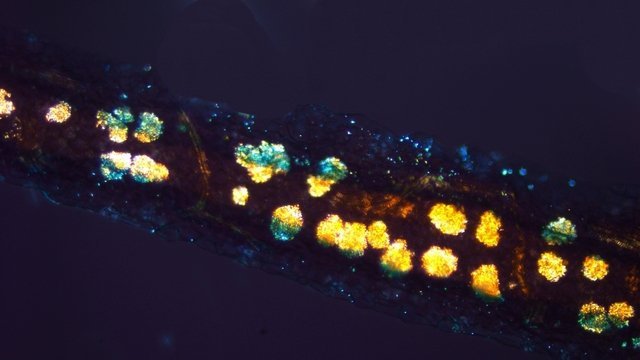
Entomosporiose feuille de cognassier Entomosporium maculatum L’ unique cognassier du jardin voit ses feuilles se couvrir tous les ans de petites taches brunes qui vont se développer en plaques mais rarement détruire toute la feuille ; les fruits sont aussi atteints mais grossissent correctement . Examen à la loupe : Regard à la loupe de la face inférieure: A noter l’importance des poils sur la face inférieure des feuilles qui persistent sur les feuilles malades. Photo de l’organisation de cette feuille - coupe du limbe – On retrouve la structure d’une feuille normale. A cuticule. B parenchyme palissadique C parenchyme lacuneux. D cellules épithéliales avec les stomates. Si on réalise une coupe de la zone atteinte par Entomosporium on constate une profonde altération de l’ architecture cellulaire. A modification des cellules de la zone palissadique. B modification des cellules de la zone lacuneuse. A hyphes courant dans le parenchyme. B début de formation d’une acervule . Disparition des deux types de parenchymes qui sont remplacés par une accumulation de petites cellules identiques ( rôles des différents enzymes synthétisés par le fungus ) Il n’existe plus de plan Les éléments cellulaires se sont développés dans une totale anarchie Conséquence de ces altérations : la photo synthèse est devenue impossible . Sur la face supérieure des feuilles sont constatées de curieuses formations plus ou moins en forme de bol les acervules : Acervule Les acervules sont des Sporophores, chez les champignons inférieurs phytopathogènes,. La production de conidies se fait dans une structure le plus souvent surélevée. C’est au milieu de ces formations que se développent les conidies ( spores ) : Pour confirmation : Extrait de l’ouvrage Microfungi of the land plants de Martin et Pamela Ellis . La mise en évidence de ces spores nécessite de placer un morceau de feuille atteinte sur une lame à cuvette et d’y ajouter de l’ eau puis on couvre avec un lamelle - il faut attendre une nuit . Sans rien toucher l’exploration du liquide autour de l’échantillon permet de découvrir quelques spores. Classification : L’agent causal de l'entomosporiose, une maladie qui affecte de nombreux végétaux, notamment des Rosacées comme le cognacier, le poirier, le pommier, le photinia, l'aubépine, etc. Classification taxonomique: Domaine: Eukaryota (Eucaryotes) Règne: Fungi (Champignons) Phylum (Embranchement): Ascomycota (Ascomycètes) Sous-Phylum: Pezizomycotina Classe: Leotiomycetes Sous-classe: Leotiomycetidae Ordre: Helotiales Famille: Dermateaceae (ou parfois Drepanopezizaceae selon les classifications plus spécifiques) Genre: Entomosporium Lév. Il est à noter que le genre Entomosporium est parfois considéré comme un synonyme taxonomique de Diplocarpon, et que l'espèce la plus couramment associée à l'entomosporiose est Diplocarpon mespili (anciennement Entomosporium mespili ou Entomosporium maculatum). Au cours de l’ exploration des échantillons on constate que ce champignon peut affaiblir l’organisme attaqué; Ainsi d’ autres champignons peuvent s’ installer - Je n’ai pas trouvé le nom de ce dernier fongus Dominique
-
Étude histologique de la rhubarbe en 3 épisodes: Inflorescence https://forum.MikrOscOpia.com/topic/20144-rhubarbe-au-microscope-l%E2%80%99-inflorescence/#comment-83679 Feuille https://forum.MikrOscOpia.com/topic/20154-rhubarbe-la-feuille/ Rhizome https://forum.MikrOscOpia.com/topic/20156-rhubarbe-le-rhizome/
-
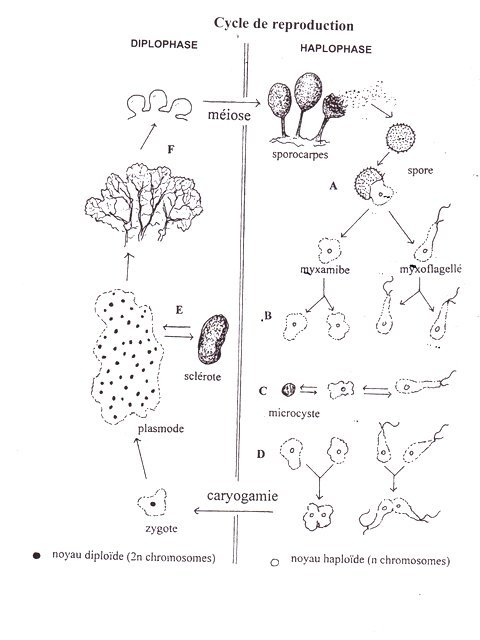
Myxomycètes 2 ( ce sujet à déjà été abordé sur MikrOscOpia ) 13 mai 2025 dans le jardin - sur une zone enrichie en feuilles un désherbage manuel est réalisé. On ne note rien de particulier .Durant la nuit il pleut,- Le lendemain est apparue sur la zone désherbée une forme que l’ on a déjà rencontrée sur MikrOscOpia : celui d’un myxomycète Fuligo septica Cette forme fait 20 cm sur 10 cm Elle est nommée un Aethalium . Le surlendemain à 20 cm du premier amas apparaît une petite plaque jaune qui va s’étendre sur 12 heures . On constate -24 heures après qu’ elle a pris un aspect brun avec quelques zones d’un jaune lumineux qui à leur tour vont se teindre en quelques heures .Le développement de cette seconde plaque s’est fait aussi dans la nuit. Elle est de la même surface que le premier amas autour de 20cm sur 10 cm. 4 photos : une toutes les 12 heures environ . Donc en un peu plus de 48 heures le myxomycète est apparu, s’est développé et est passé d’un développement végétatif à une sporogenése. Le plasmode : Il s’agit ici d’une masse souple jaune lumineuse qui s’est déplacée ici sur 20 cm en quelques heures sur un terrain humide Les limites semblent faites d’un film biologique plus que d’une membrane constituée – Son extension a été de type amiboïde : développement de prolongements digitiformes qui se sont épaissis rapidement . La constitution interne est un cytoplasme .Il y est trouvé un nombre considérable de formations rondes de très petite taille 1,6 µm En fait il s’ agit de granules calcaires ( disparition s’il est ajouté 1 gtt d’ acide chlorique dilué au 10%). .Ces granules vont jouer un rôle très important - voir plus loin. Au milieu de cette prolifération de microgranules( B ) Il existe des cellules plus volumineuses de couleur orangée qui pourraient être des noyaux ( A) . La répartition des diverses structures présentes n’est pas homogène. Elle semble suivre des lignes mais il n’ y a aucune cloison ( certaines publications parlent de la présence de veines où s’écoule le cytoplasme avec des mouvements alternatifs Ce qui n’ a pas été observé ici : intérêt dans ce cas du microscope électronique )……. Dans cette observation le cytoplasme baigne l’ensemble des structures ( microgranules + noyaux ( la multiplicité des noyaux dans une même cellule est appelée un Syncitium). Le plasmode est donc une masse visqueuse multi nucléée qui se déplace en rampant sur les surfaces à la recherche de nourriture (bactéries, spores de champignons, matière organique)..L’ absorption se ferait sur le mode phagocytose. La digestion est probablement faite directement grâce aux enzymes contenus dans le cytoplasme .L’eau est probablement absorbée par osmose ce qui explique la grande fragilité des myxomycètes à la sécheresse des supports .Sécheresse qui induit un passage en forme de résistance du myxomycète. . A ce moment de l’évolution il y a une métamorphose de la structure du plasmode sur un temps très court - L’ observation du plasmode constate que sous l’ effet du vent et du soleil des petites taches brunes apparaissent qui vont recouvrir la totalité de la surface Le Myxomycète est devenu ferme avec une surface légèrement granulée et une masse franchement déshydratée .Il a pris la forme d’un Sclérote . Cette forme de résistance a pris le nom de Sclérote par assimilation car cette dénomination est plutôt réservée aux champignons dans leur forme de résistance. Cette transformation n’a pas pris plus de 8 heures. Une coupe met en évidence la nouvelle structure interne Il n’y a plus de cytoplasme tout est transformé en une masse brun sombre hétérogène. A Peridium qui forme une croute rêche et brune B Capillitium qui correspondrait à la fusion d’un grand nombre de Sporocystes ( mais ici ces structures n’ont pas été mises en évidence ) cette sporothéque est maintenue par en ensemble de membranes indurées qui fragmentent le contenu. C Hypothale qui forme le pied de l’ Aethalium et qui s’ emmêle avec les débris de terre et de feuilles sur lesquels repose le Myxomycète( repose car cette structure ne possède pas de rhizoïde donc ne pénètre pas le milieu externe). Image du Peridium Cette zone de couverture superficielle est faite de petites plaques sèches et dures et doit correspondre au résultat de l’asséchement du cytoplasme associé aux granules calcaires ( l’ équivalent d’un ciment ). Si on dissocie cette plaque : on constate l’image suivante: On constate qu’ il n’ existe aucun élément cellulaire .mais un amas de matériel sec.disposé en plaques ondulées .sur plusieurs couches. Image du capillitium ( B ) 2 éléments sont visibles : des plages noires qui correspondent à l’accumulation des spores et des travées blanches de renforcement qui organisent l’ensemble .du capillitum Il y a des millions de spores. Image des structures blanches de renforcement. Il n’y a pas d’élément de type fibreux mais des éléments semblables à de la colle durcie -( constitués probablement d’une coagulation du cytoplasme + granules calcaires ) En fait en dehors des spores et des noyaux il n’ existe aucun élément de type cellulaire. Image de l’ hypothale Cette zone est la zone sur laquelle repose la plasmode .Au stade initial cette zone est fluide et visqueuse favorisant le glissement ;Comme le reste de la structure cette zone s’est durcie ,engainant des éléments du terrain,.lors de la transformation en Sclérote. ( A ). Là encore l’ aspect histologique est acellulaire correspondant à un durcissement du cytoplasme mais de manière beaucoup plus souple ,plus humide qu’ au niveau du Péridium . La couleur n’ a pas changé elle reste cependant d’un jaune moins brillant que le jaune du plasmode. La suite Un morceau de Aethelium est placé en milieu humide dans l’espoir de mettre en évidence des myxamibes ou des myxoflagelés. Mais on se trouve devant une difficulté : le milieu est très habité par un tas de petits protistes et il est difficile de savoir qui est qui ( il faudrait utiliser les techniques d’ isolement ) Schéma du cycle de reproduction référence 1 En un sens, on peut dire que le plasmode d'un myxomycète se comporte comme s’ il était formé d'une seule cellule géante. Voici pourquoi : Syncytium : Le plasmode est un syncytium, ce qui signifie qu'il s'agit d'une masse cytoplasmique unique contenant de nombreux noyaux. Contrairement à nos cellules individuelles qui ont un seul noyau, le plasmode est une sorte de "super-cellule" avec une multitude de centres de contrôle génétique flottant librement dans le même cytoplasme. Mouvement coordonné : Malgré la présence de nombreux noyaux, le plasmode se déplace et se nourrit de manière coordonnée. L'ensemble de la masse protoplasmique flue et se déplace en réponse aux stimuli, comme si une seule entité prenait des décisions. Réponse unifiée : Lorsqu'une source de nourriture est détectée, l'ensemble du plasmode se dirige vers elle. De même, face à des conditions défavorables, il peut se transformer en sclérote, une forme de résistance durcie, là encore comme un seul organisme réagissant à son environnement Ref Les Myxomycétes de Michel Poulain Marianne Meyer Jean Bozonnet edité par la fédération mycologique et botanique Dauphiné -Savoie Dominique
-
Exploration d'un bac extérieur rempli d' eau
Dominique. a répondu à un(e) sujet de Dominique. dans ÉCOSYSTÈMES MICROSCOPIQUES
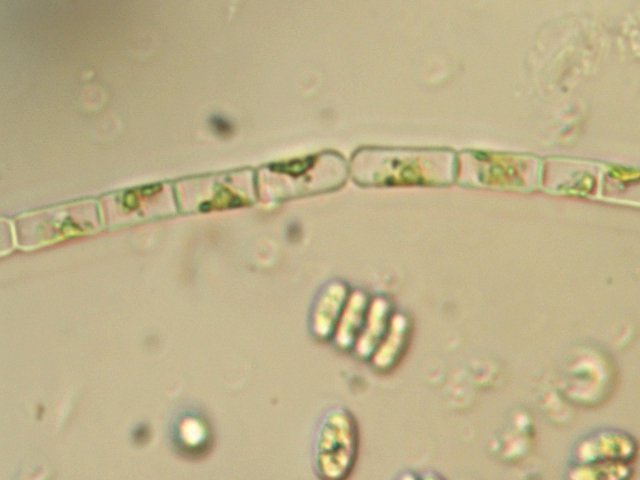
Bonsoir Jean-Marie Geminella me semble la bonne piste mais The fresh water alga oriente vers une autre Chlorophyta Sur la photo on remarque qu'il y a un enduit gélatineux épais tout le long de l 'algue ( A peine visible mais bien présent ) il semble donc s 'agir de Gloeotila monospora ( cell within a wide mucilaginous envelope and empty cells ) Merci de ton aide Amicalement Dominique -
Exploration d'un bac extérieur rempli d' eau
Dominique. a répondu à un(e) sujet de Dominique. dans ÉCOSYSTÈMES MICROSCOPIQUES
Bonjour i ll te faut acquérir deux éléments un micromètre oculaire et un micromètre objet . On peut en trouver sur Amazone mais il faut que le micromètre oculaire s' adapte à l 'oculaire de ton microscope ( donc voir le fabriquant). Amicalement Dominique -
Exploration d'un bac extérieur rempli d' eau
Dominique. a posté un sujet dans ÉCOSYSTÈMES MICROSCOPIQUES
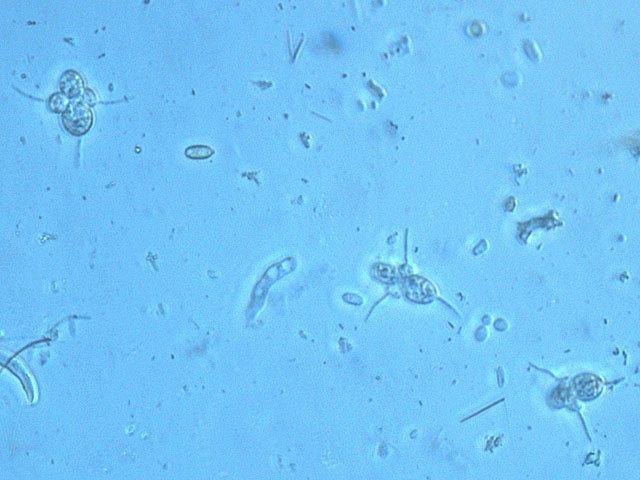
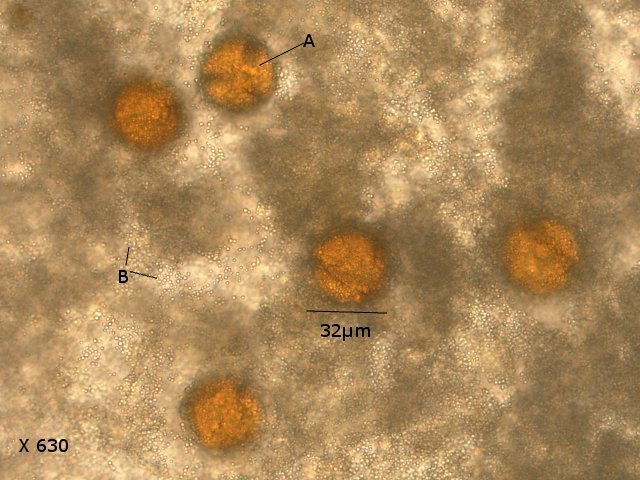
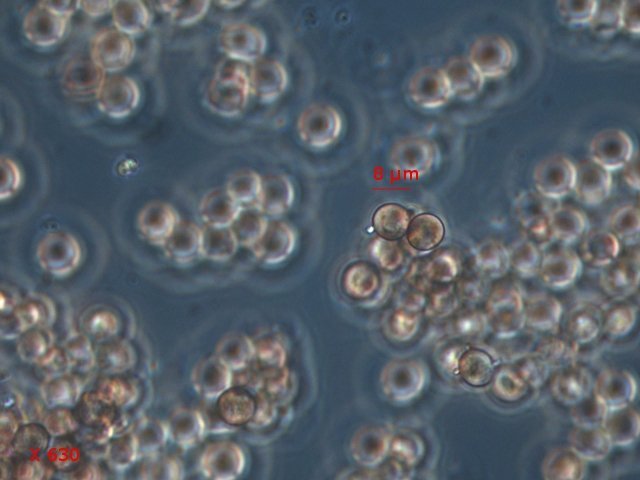
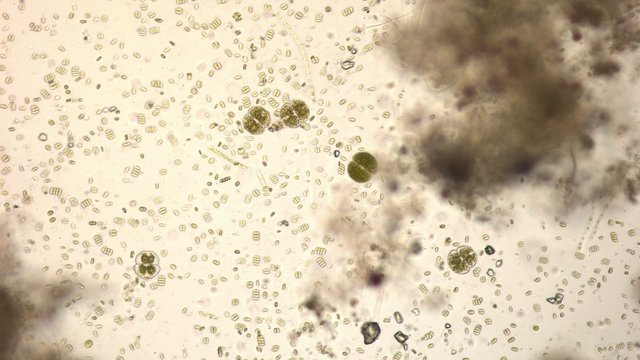
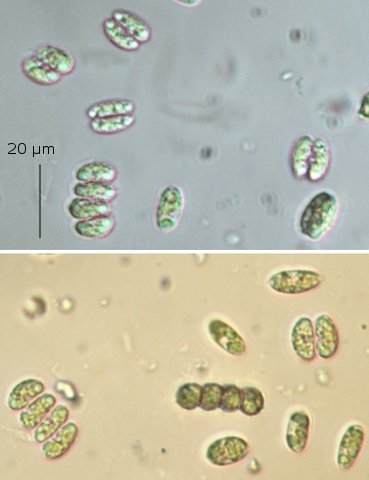
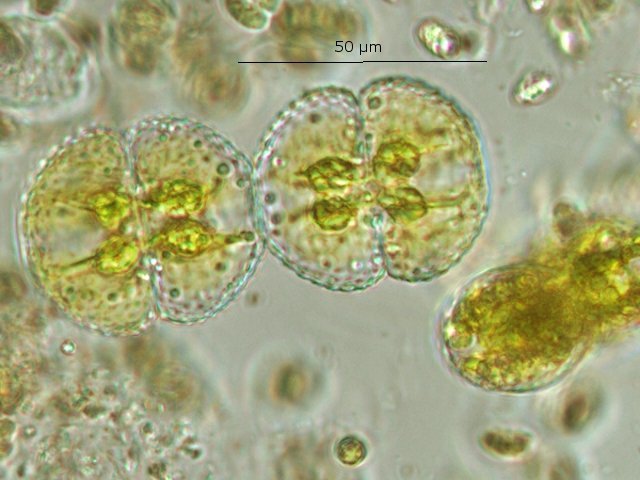
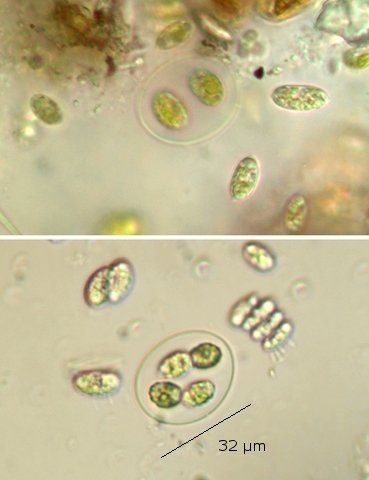
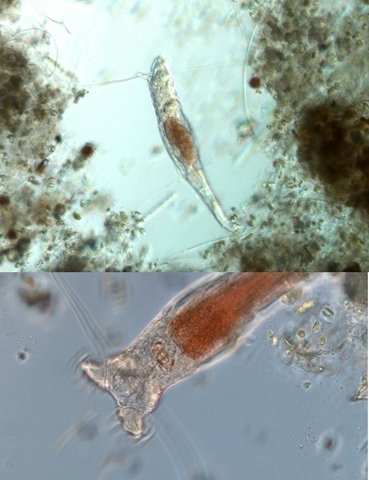
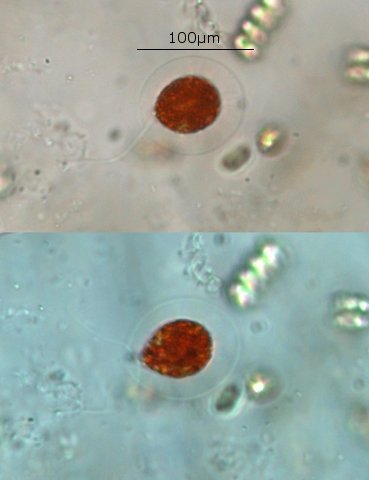
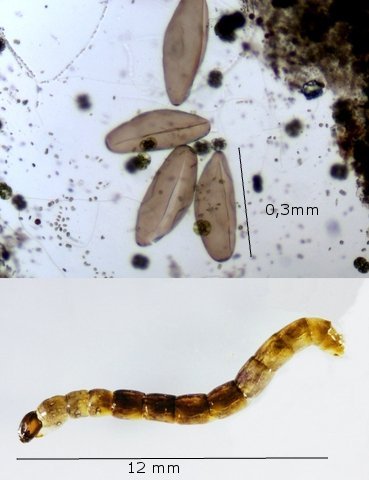
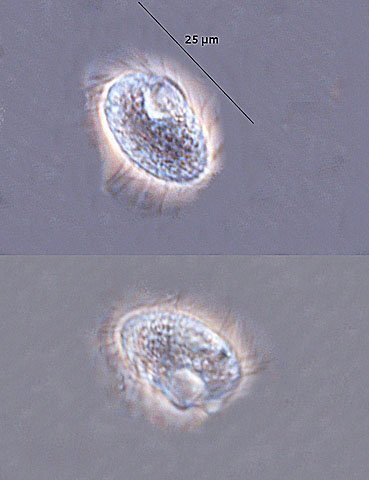
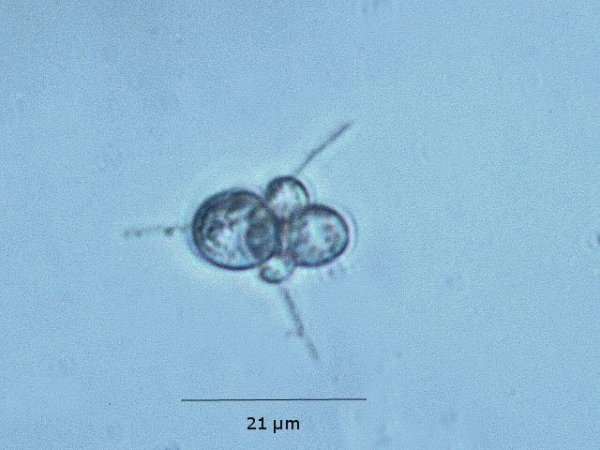
Exploration d'un bac extérieur rempli d' eau L’ histoire remonte à l’année dernière . je place un bac vide à l’ extérieur de la maison - sur le sommet d’un petit muret. L’évolution a été très lente , le bac s’ est doucement rempli et très rapidement - de l’ eau est devenue verdâtre et trouble . C’est en fin aout que l’ on constate la présence de larves de chironomes . Durant l’ automne quelques feuilles sont tombées dedans apportées par le vent - elle y sont restées tout l’ hiver sans grand changement . L’ eau du bac est redevenue parfaitement transparente durant l hiver et ses températures froides. Par contre elle s’est reverdie avec l’ arrivée du printemps - Les feuilles se sont mises à changer d’ aspect - Le limbe des feuilles a disparu en moins de 1mois et des amas d’ une substance hétérogène et aérée sont apparus formant de petits amas .Ces amas surnagent à la surface du bac . Les nervures de la feuille sont très apparentes dépourvues du parenchyme foliaire qui a été détruit par l' activité microbienne Coloration Acridine / Bleu Alcian Les fibres des vaisseaux sont dénudées et leur armature spiralée est bien visible . L’examen des amas surnageant met en évidence une flore et une faune très diversifiée. Il existe des algues du genre Desmodesmus - Il en existe au moins deux espèces: Présence d’une famille de Cosmarium – les parois sont granulées les demi –cellules sont subsemiciculaires pouvant correspondre à Cosmarium Botrytis. Il y a aussi des algues filamenteuses ( introuvables dans mes catalogues en particulier parmi les cyanobactéries). L’ algue suivante ressemble beaucoup à Glaucocystis nostochinearum: Il existe aussi des rotifères Bdelloïdes en grande quantité apparemment de même espèce .Ils sont difficiles à mesurer leur taille est autour de 160 – 320 µm . Les nombreuses taches rouges correspondent à la multiplication des Haematoccus qui avec leurs flagelles peuvent se déplacer très rapidement. Il est aussi retrouvé des enveloppes ouvertes de type œufs -L’ hypothèse est qu’ il s’agit des œufs des chironomes -Ces chironomes avaient presque disparu durant l’ hiver pour réapparaitre il y a un mois - en réalité il y a toujours eu 1 à 2 petits diptères sur le bord du bac ( ce qui veut dire que le froid de l’ eau a ralenti les métamorphoses sans les arrêter ) . Enfin il est aussi trouvé la présence d’un petit nombre de ciliés La taille de ce cilié est petite et pourrait correspondre avec la classe des Cyrtophoria espèce Chilodon mais sans aucune certitude ( dans les catalogues les ciliés se ressemblent terriblement ). Sans oublier l’ énorme population bactériennes en grande partie responsable de la dégradation des feuilles et de l’ enrichissement du milieu . Il existe un élément intriguant : la multiplication des micro filaments dans tout le milieu. En conclusion : Partant d’une cuve d’eau de pluie il s’est développé en 1 an1/2 un micro monde . D’où vient l’ensemencement :la pluie - le vent - les feuilles - et les insectes volants ont fait leur travail de propagation des formes vivantes l' expériences va se continuer encore sur 1 an. Dominique.
-
Population de l'estran de la plage du Havre
Dominique. a répondu à un(e) sujet de Dominique Voisin dans ÉCOSYSTÈMES MICROSCOPIQUES
Bonjour Dominique comment fais tu pour obtenir tes échantillons ? - Jean -Marie conseille de faire une filtration sur un tissus à mails très fines d'un prélèvement d'eau cela plusieurs fois de suite pour augmenter ses chances ou utilises tu un filet à plancton ? On voit que ta pêche est bonne et diversifiée. Amicalement Dominique -
Petroneis monilifera
Dominique. a répondu à un(e) sujet de Dominique Voisin dans Diatomées (Collections)
Bonsoir Dominique Le fait de ne pas faire subir aux diatomées les préparations classiques fait que l' on du mal à les reconnaître quand aucune préparation n' est réalisée .Je trouve qu'il y a autant de plaisir à les regarder comme tu nous les présentent ici qu' a admirer leur squelette décharné. Amicalement Dominique. -
Talle chez certaines poacées Blé - Orge - Plateau de tallage
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
Bonsoir Solito de solis Tu as raison des céréales comme le riz , le maïs , le quinoa,le sarrasin , l'avoine peuvent remplacer le blé .En ajoutant des pommes de terre; des patates douces et d' autres légumineuses on obtient la ration correcte en glucides , en fibres , en vitamines et en minéraux Mais l' avantage avec le blé est qu il est de culture facile sous beaucoup de climats et qu'il permet d' avoir des rendements importants pouvant faire face à l' alimentation des 8 milliards d' humain et selon les prévisions 10 milliards futures - car le fond du problème tient dans ces chiffres - personne ne sait en l état actuel de nos possibilités , qui se dégradent , qu elle va être la solution si elle existe. Amicalement Dominique. -
Talle chez certaines poacées Blé - Orge - Plateau de tallage
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
Bonjour Jean- Luc Merci pour ta réponse . Ce petit article sur la talle du blé et de l’orge est en fait une introduction au virage que notre société va devoir faire face . La situation pour le future est en effet sombre -les récoltes ne peuvent plus dépasser les limites atteintes à ce jour et cela depuis plus de 2O ans. -les surfaces des terres agricoles se réduisent partout dans le monde ( urbanisation ou disparition naturelle ) et la qualité de celle qui restent cultivables est de moins en moins bonne. --les conditions climatiques se détériorent plus vite que la capacité d’ adaptation des plantes cultivées ne le font . --En Europe se développe le biologique et donc l’ interdiction de certains intrants ou phytosanitaires indispensables pour obtenir des rendements élevés. Ces rendements élevés sont cependant indispensables à maintenir en raison d’une croissance apparemment inexorable de la population. Le Kwashiorkor ne fait pas encore partie des maladies rencontrées en Europe mais au train où vont les choses d’ici pas très longtemps il se pourrait que cela deviennent réalité . Amicalement Dominique -
Talle chez certaines poacées Blé - Orge - Plateau de tallage
Dominique. a posté un sujet dans Dominique
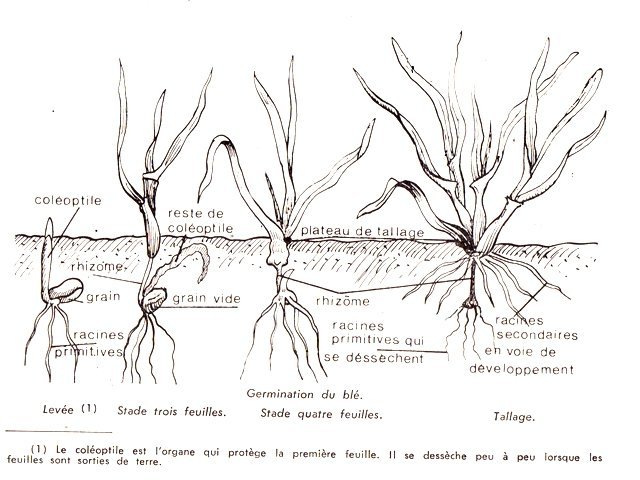
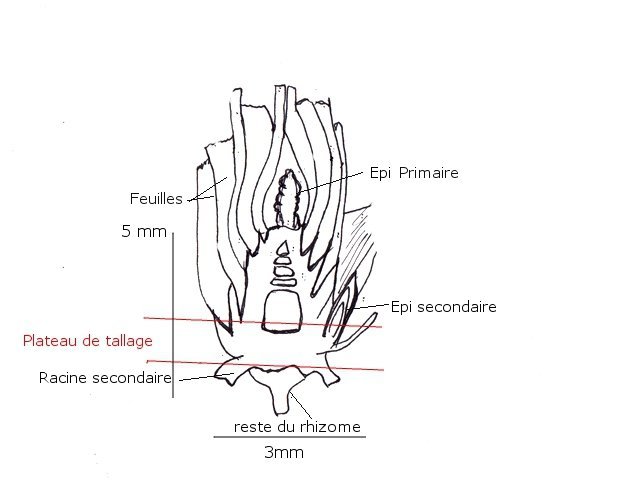
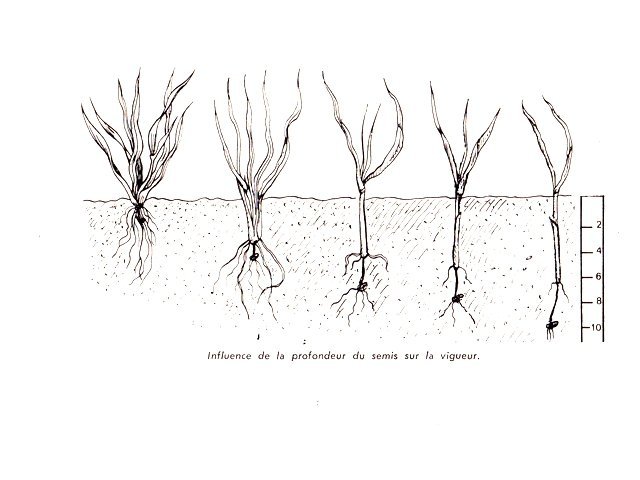
Talle chez certaines poacées Blé - Orge Les semis ont été faits en fin d’année - Du fait des températures la germination des grains s’ est faite doucement – mais avec l’élévation des températures le développement de la seconde phase de la croissance a commencé Aspect ( le 2 Mars ) actuel des plantules : Photo du collet de la plantule Avant de commencer ce petit article il faut donner une précision orthographique Thalle ( avec un H masculin ) Définition : Un thalle est un appareil végétatif simple, non différencié en racines, tiges et feuilles. On le trouve chez les organismes tels que les algues, les champignons, les lichens et certaines plantes primitives comme les mousses et les hépatiques. Caractéristiques : Il n'a pas de tissu vasculaire (conducteur de sève). Il peut avoir des formes variées : filamenteuses, laminaires, crustacées, etc. Il assure les fonctions de nutrition, de croissance et de reproduction. Exemples : Le corps d'une algue marine, le mycélium d'un champignon, le corps d'un lichen. Talle ( sans H féminin) : Une talle est une tige secondaire qui se développe à partir de la base d'une plante, notamment chez les graminées (Poacées). Le processus de production de talles est appelé tallage. Caractéristiques : Elle se forme à partir du plateau de tallage, la base de la plante. Elle porte des feuilles et peut produire des inflorescences (épis, panicules, etc.). Le tallage permet à la plante de se ramifier et d'augmenter sa production de graines. ! L'orge et le blé, comme la plupart des céréales, ne développent pas de rhizome au niveau du plateau de tallage. Elle appartient à la famille des Poacées (graminées) et se caractérise par un système racinaire fasciculé. Schéma du développement de ces deux graminées: On voit sur ces schémas le nom de rhizome qui est le nom donné à la première racine issu du grain – mais ici il n’ a aucun rôle dans le développement des structures secondaires et cette ébauche embryonnaire disparait ensuite. Organisation générale d’une talle: Le plateau de tallage a une épaisseur de 7 à 10 mm Il est divisible en deux parties la partie supérieure où les cellules vont se différentier en epis secondaires La partie inférieure où les cellules vont se différentier en racines secondaires Si au départ ce plateau n’ est formé que d’une talle ( la talle primaire ) au fur et à mesure du développement le plateau sera le résultat de l’ accolement de plusieurs talles. L’ extension du plateau de tallage se fera de manière centrifuge les racines et les épis nouveaux qui vont se developer le font toujours sur la periphérie du plateau de tallage Aspect macroscopique: A tige principale a tiges secondaires b racines secondaires Coupe d ' une talle. : Image d’ une talle unique A partie initiale de la tige principale B zone des méristémes C nouvelle racine qui se developpe en peripherie ( on parle de systéme racinaire fasciculé ) D restes des racines primaires et du rhizome qui ont disparus Le tissu qui constitue la zone des méristèmes est très hétérogène et ne présente pas d’organisation bien définie il constitue la zone de fabrication d’ un bourgeon d’ où naitra la prochaine talle. ( X 60 ) Pourtant au niveau programmation la partie supérieure exprimera des tiges et la partie inférieure exprimera des racines A zone d’ expression des tiges B zone d expression des racines C zone residuelle de la talle primitive. A tige primaire. B deux tiges secondaires. Cette coupe montre l’ accolement de plusieurs talles (A) soit le départ des tiges florifères des différentes talles secondaires ces talles sont toutes issues des bourgeons axillaires de la premiere talle ,Les bourgeons axillaires des premières talles peuvent à leur tour se développer : apparaissent alors des talles tertiaires. Une touffe peut donc être vue comme une colonie clonale à extension périphérique (Le tallage est recherché dans le cas des gazons pour assurer la formation d'un tapis de végétation dense.) En résumé : Le tallage est caractérisé par l'entrée en croissance de bourgeons différenciés à l'aisselle de chacune des premières feuilles. Le développement de ces bourgeons au niveau des nœuds donne naissance aux talles primaires. Chaque talle primaire émet des talles secondaires susceptibles d'émettre des talles tertiaires : l'aptitude à émettre en plus ou moins grand nombre des talles secondaires et tertiaires est une caractéristique spécifique et variétale. Les racines définitives de la plante se forment à partir du plateau de tallage qui soutient le maître-brin et les talles. Comme pour les feuilles, chaque talle apparaît après un intervalle de temps constant, appelé phyllochrone. Chez les céréales, la durée du phyllochrone peut varier entre la tige principale et les talles. Une jeune talle dépend de la pousse parentale pour la photo-assimilation jusqu'à ce qu'elle ait développé plusieurs feuilles et un système racinaire adéquat. Bien qu'une talle mature puisse sembler fonctionner comme une entité indépendante, il existe apparemment une relation entre les talles interconnectées par un système vasculaire commun. Ainsi, une graminée semble être un système hautement organisé plutôt qu'un ensemble de talles concurrentes. Origine des talles Au niveau de l’embryon dans la graine, l’apex au stade végétatif émet successivement des structures, les phytomères, composées systématiquement de trois éléments : l’ébauche d’une feuille, un nœud et un bourgeon axillaire. Entre la levée et la transition florale, la tige principale d’un pied de blé, appelée brin maître, va constituer dix à quinze phytomères végétatifs, et donc autant de feuilles et de bourgeons axillaires, cachés le long de la tige à l’aisselle de la feuille (figure 1). Ce sont les bougeons des premières feuilles qui, en sortant de leur dormance et en poussant à leur tour, vont produire les talles. Ces talles sont elles-mêmes, à l’image du brin maître, constituées de phytomères qui contiennent des feuilles, des nœuds, et des bourgeons. Donc le processus peut se reproduire en périphérie de la nouvelle talle. Comment favoriser le tallage donc le rendement des cultures A- La profondeur du semi . B la richesse en azote du milieu Ainsi, l’azote n’accélère pas le rythme d’apparition des talles, mais il augmente la probabilité qu’une talle soit effectivement émise Cette richesse en azote va dépendre aussi de l’hydromorphie de la terre ( l’eau dilue l’ azote ) C dans le cas de sécheresse , le tallage herbacé peut être très fortement réduit, et irrémédiablement pénalisant pour l’élaboration du rendement. D l’espace libre entre deux plantes ( donc la densité du semis) Plus la densité de végétation est élevée, plus l’émission des talles sera inhibée. Une telle inhibition s’observe notamment dans des parcelles à forte densité de végétation, ou à pression adventice élevée : les plantes émettent alors moins de talles E le roulage si la terre le permet ( pas trop humide ) le roulage favorise le développement des talles et permet de réduite la quantité d’azote à apporter Conclusion Le tallage du blé et de l’orge est un des points clefs du rendement de ces céréales. En 1950 le rendement moyen était de20 quintaux à l’ hectare en 2024 70 quintaux en moyenne . Mais les rendements stagnent désormais Le ralentissement de cette progression, voire ses irrégularités constatées semblent reliés à une certaine désintensification résultant d’une économie d'intrants (réduction de 15-20 % des apports azotés, des fongicides, moindre travail du sol...) et du changement climatique (fortes températures au remplissage du grain, faible luminosité, pluviométrie excessive...). On doit s’interroger sur l’impact que pourrait avoir sur la productivité un abandon des variétés de blé modernes et des produits de synthèse, ce qui pourrait conduire rapidement à rendre notre pays dépendant des importations, situation qui prévalait avant 1950. Références : ref https://botarela.fr/Poaceae/Description-detail/Tige-et-feuilles.html ref : https://www.perspectives-agricoles.com/recherche-agronomie/le-tallage-une-cle-de-la-productivite-du-ble Dominique
-
-
Cochenille les dégâts sur les branches .
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
Bonsoir Jean-Luc Merci pour ton article qui va dans le sens d'une transmission virale responsable d une atteinte du Liber .Les coupes effectuées dans cette observation montre que c'est le liber qui est atteint - donc virus ? toxique? - En tant qu'amateur on n'a ni la connaissance ni le matériel pour avancer ,, dommage.......... Bien amicalement Dominique.





























.jpg.511b6b26d6175129027a72fab821e6c0.jpg)