Dominique.
-
Compteur de contenus
1 474 -
Inscription
-
Dernière visite
-
Jours gagnés
1
Tout ce qui a été posté par Dominique.
-

Cochenille les dégâts sur les branches .
Dominique. a répondu à un(e) sujet de Dominique. dans Dominique
Bonsoir Jean-Marie Ton hypothèse d'une infection bactérienne ou viral transmise par la cochenille est je pense une bonne hypothèse .Pour la cochenille il faudra trouver des travaux s 'ils existent . Dans le livre Cultiver et soigner les arbres édition Eyrolles les insectes sont une des grandes causes à de nombreuses maladies .Mais le lien entre insectes et lésions créées ne sont pas aborder. c'est dommage. Amicalement Dominique. -
Cochenilles : atteinte sur les bourgeons
Dominique. a posté une image de la galerie dans Galerie Mikroscopia
-
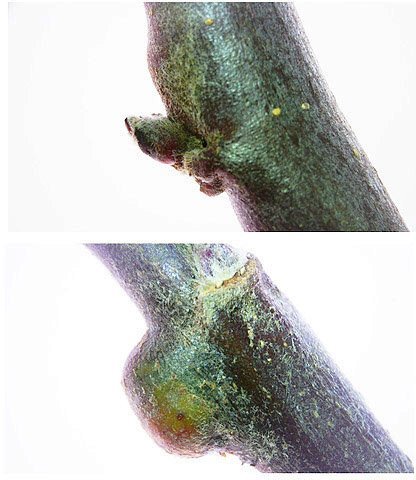
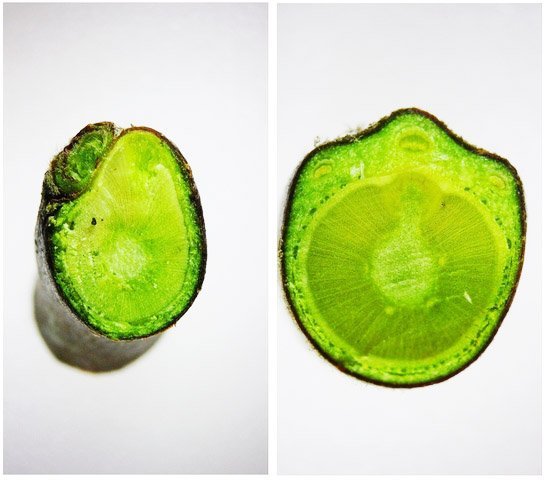
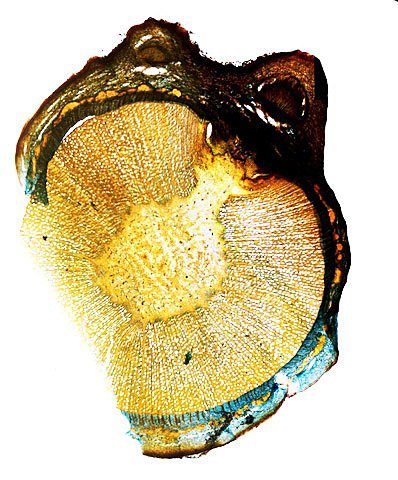
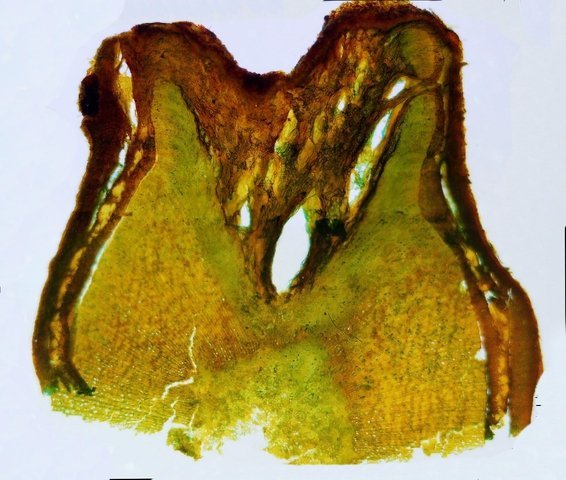
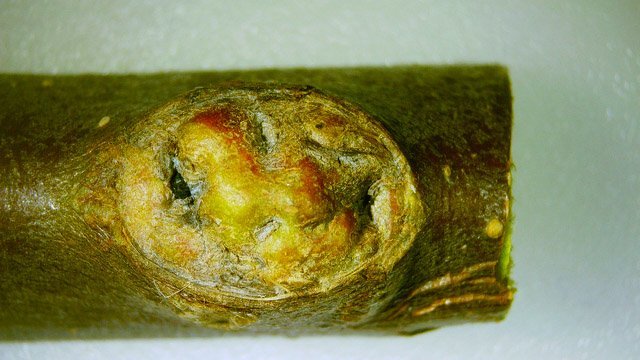
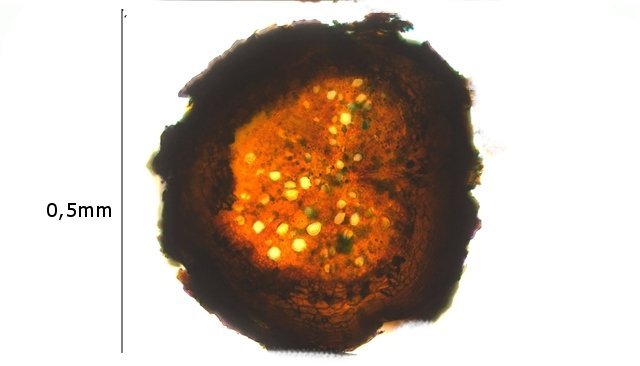
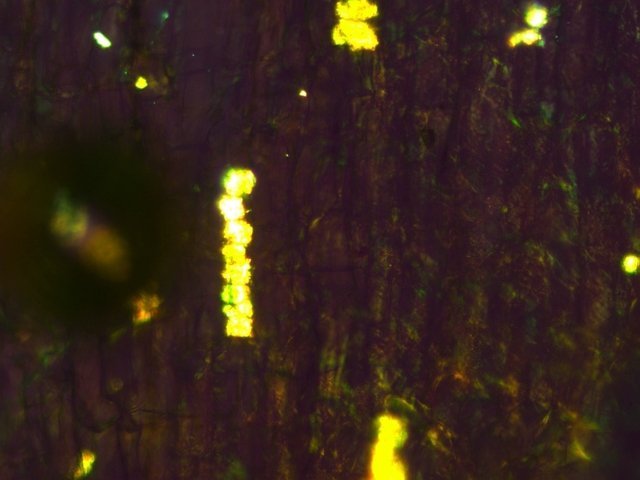
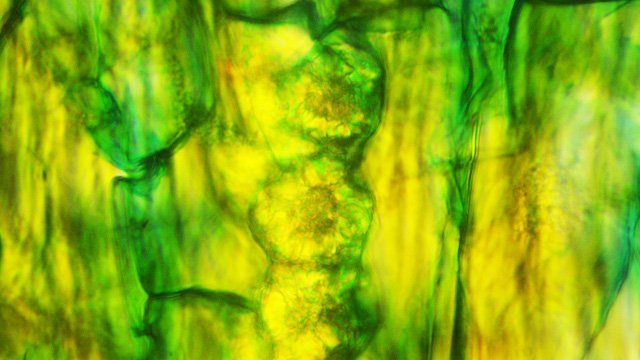
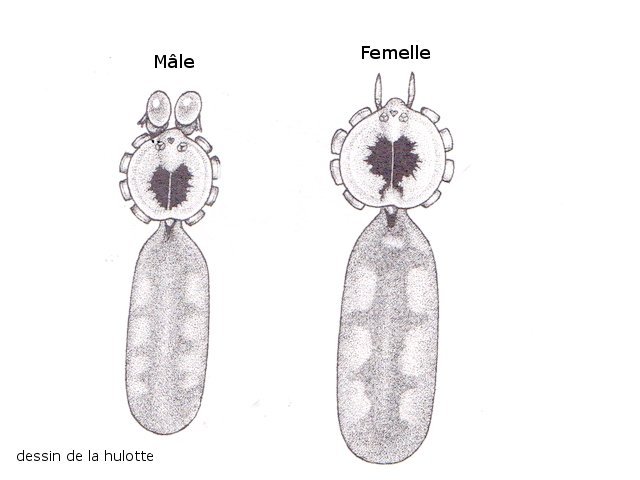
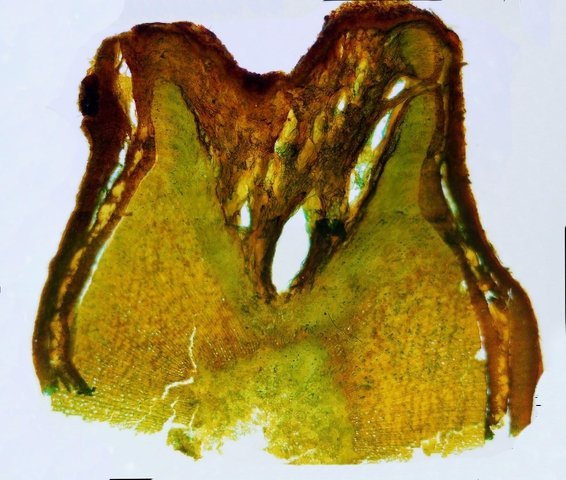
Cochenille les dégâts sur les branches des arbres. C’est la période de la taille des arbres fruitiers - Sur un des pommiers il avait été noté durant l’ été la présence de cochenilles .Cet hiver on constate des déformations sur les branches de l’ année La cochenille passe l’hiver au stade larvaire, soit dans le sol, soit cachée dans les structures d’ une serre. Elle émerge dès que les températures s’élèvent et en présence de plantes hôtes. La cochenille a déjà été présentée sur MikrOscOpia https://forum.MikrOscOpia.com/topic/20038-cochenille/#comment-83036 Sur cette observation le rostre de l’ insecte est très bien mis en évidence – ce rostre permet à l’ insecte le prélèvement de la sève élaborée des arbres sur lequel il repose ( donc prélèvement dans le liber ) Sur cette observation la cochenille responsable semble être de la famille des : Pseudococcidées, Pseudococcidae Chez cette famille d'Homoptères on rencontre les fameuses cochenilles farineuses, Pseudococcus spp. et Planococcus spp. Ces insectes ont la particularité d'être recouverts de filaments blancs et cireux. ***************************** La présence indirecte de cochenilles est repérable dés l’ hiver –les rameaux de l’ année sont porteurs de bourgeons qui ont totalement changé de forme - Les rameaux sont couverts de boursouflures ( ces boursouflures vont persister sur les branches plus anciennes). L’ennui est que l’émergence des feuilles ne va pas se faire sur ces bourgeons altérés. ;( elles peuvent aussi se faire sur un mode atrophique déformé) Observation faite uniquement avec le microscope à écran Tomlov Comparaison entre un bourgeon sain et un bourgeon transformé: Sur certaines boursouflures on repaire encore la présence de filaments blancs -restes de cochenilles ou poils du bourgeon du pommier ? La question posée dans cette présentation : n Quelles sont les modifications de l’architecture ? n Et une question sans réponse ? - qu’elles sont les produits contenus dans la salive des cochenilles qui peuvent entraîner un tel désordre Coupe de bourgeons foliaires en bonne santé . Aspect des dégâts sur les bourgeons : Dans le cas d’une altération superficielle - la naissance des feuilles sera possible ces feuilles ne seront pas parfaites mais resteront en partie fonctionnelles. Autres aspects sur d’autres branches de l’ atteinte des bourgeons foliaires . Ces images montrent que l’altération est plus ou moins profonde ( altérant la structure cellulaire depuis l’épithélium superficiel jusqu’ à la zone des méristèmes primaires Dans ce cas ( image ci-dessus ) l’ altération de l’ architecture est beaucoup plus prononcée et ne donnera pas de naissance à des feuilles fonctionnelles . Aspect microscopique de ces deux états : Aspect histologique des bourgeons. Problème technique La coupe fine est très difficile en raison de la dureté du bois La coloration choisie est ( Acridine ( rouge ) + Bleu Alcian ) Pour éviter les bulles d’air très abondantes sur ce type de préparation on réalise avant le montage 1 bain dans alcool isopropylique 100% puis 1 bain dans Limonéne et enfin 1 bain dans essence d’ Euparal et montage dans l Euparal Aspect d’ un bourgeon sain : Aspect histologique d’un bourgeon altéré par les piqures de cochenilles Il existe aussi des places sur la branche qui présentent des anomalies de structure en dehors de la présence de bourgeons foliaires. Et cela pour des lésions s’ étendant sur une longueur de 1à 4 cm. La salive de certaines espèces de cochenilles est toxique pour les plantes. Cette toxicité se manifeste de différentes manières : Déformations et décolorations : La salive peut provoquer des réactions anormales dans les tissus végétaux, entraînant des déformations des feuilles ou des tiges, ainsi que des changements de couleur. Inhibition de la floraison : Dans certains cas, la salive toxique peut perturber le développement des boutons floraux et empêcher la plante de fleurir. Affaiblissement général : La salive peut interférer avec la capacité de la plante à absorber les nutriments et l'eau, ce qui la rend plus vulnérable aux maladies et aux autres parasites. Ces atteintes sont variables suivant le type de cochenilles présentes Cochenilles à bouclier : Leur salive peut provoquer des taches jaunes ou brunes sur les feuilles, qui finissent par se nécroser et tomber. Cochenilles farineuses qui est le sujet de cet article : Leur salive peut entraîner ’une altération des structures végétales et favoriser le développement de fumagine, une maladie fongique qui recouvre les feuilles d'une substance noire et collante. Traitement Le traitement classique était l’ application au pulvérisateur d’ un mélange de Malathion et d’ huile de pétrole en fin d’ hiver ( mais retrait du commerce du Malathion en 2008 qui était aussi un anti poux très efficace avec le léger inconvénient de sentir transitoirement la poubelle ) . Traitement biologique contre les cochenilles, est le suivant Pulvérisez : un mélange d'huile végétale de type huile de colza ou huile de pétrole et de liquide vaisselle à parts égales, dilués à proportion d'une cuillerée à café (= 5 ml) pour 150 ml d'eau ; ou faire une pulvérisation de la solution d’ une solution de 75 g de savon noir liquide dans 1,5 l d'eau.y ajouter 150 cc d’ alcool à bruler faire 3 applications de cette solution à 3 semaines d’ intervalle Références: https://www.bio-enligne.com/homoptere/480-cochenilles.html https://www.researchgate.net/publication/313862365_MANAGEMENT_OF_COCHINEAL_Dactylopius_coccus_Costa_INSECT_PEST_THROUGH_BOTANICAL_EXTRACTION_IN_TIGRAY_NORTH_ETHIOPIA Dominique.

-
-
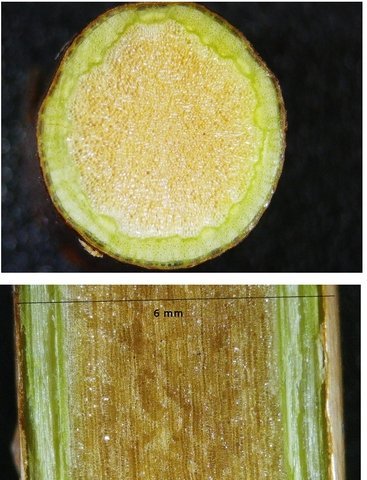
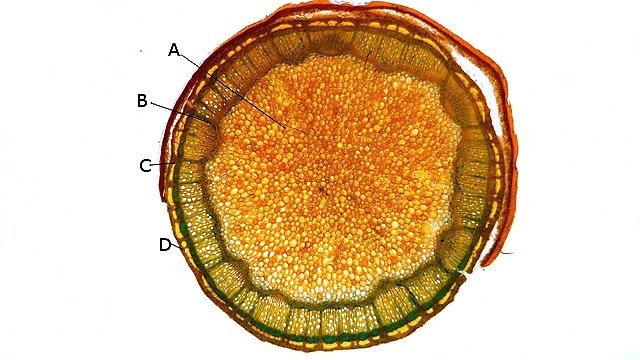
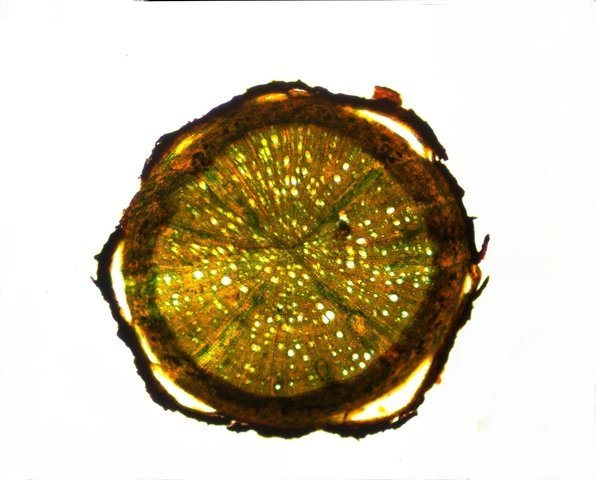
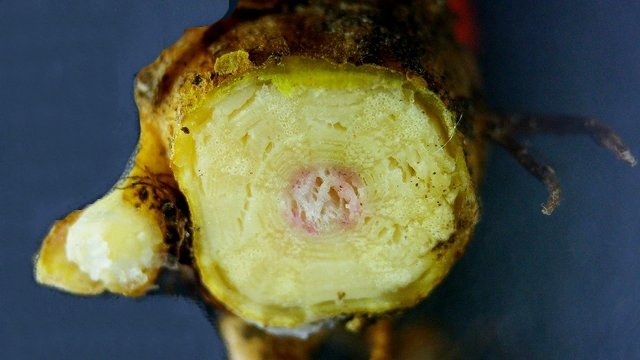
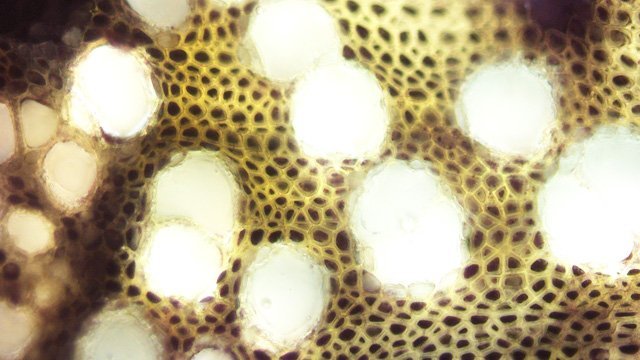
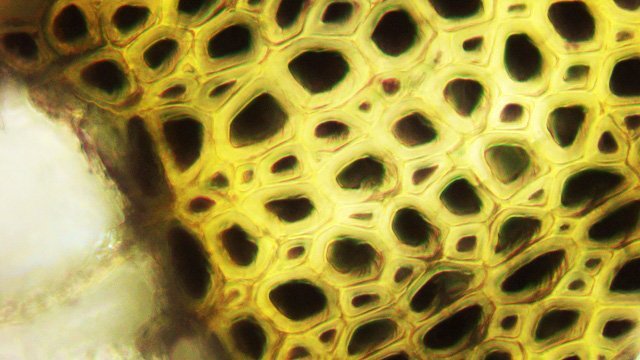
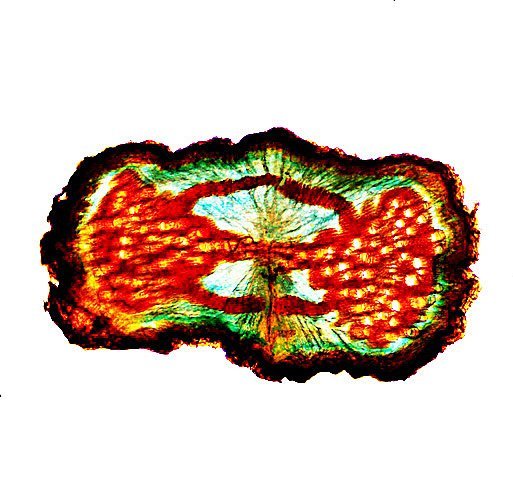
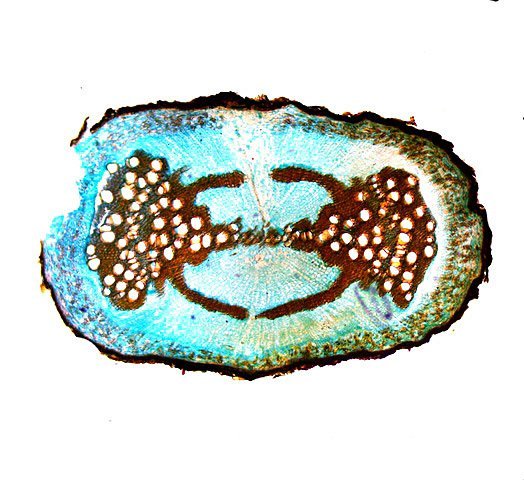
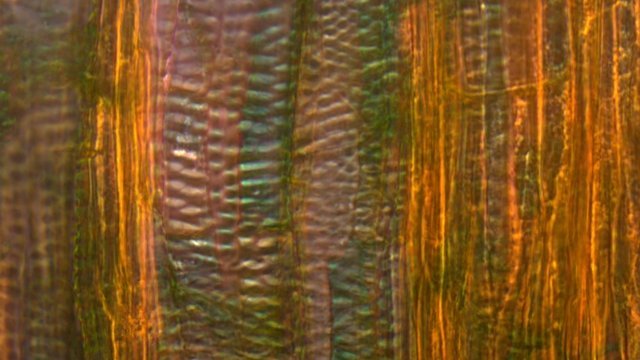
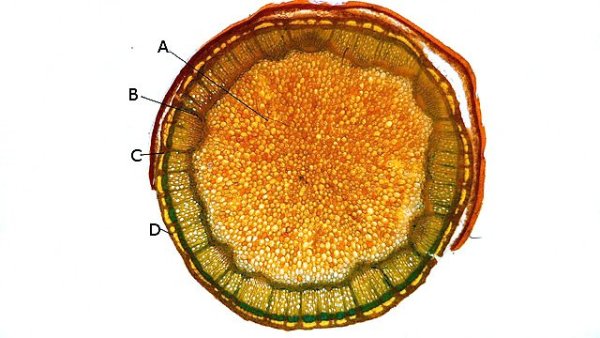
Framboisier Rhizome Le framboisier a toujours tendance à déborder le lieu qui lui a été assigné ;la vitesse de propagation est lente .Dans le jardin de jeunes pousses apparaissent au printemps 20 cm plus loin que les pieds mère. La raison est liée au développement d’un puissant rhizome Il y a un an le rhizome de la Renoué du japon qui peut atteindre plus de 20 mètres a été présenté et le mois dernier le rhizome de l’ortie Rhizome du framboisier : A --Tige aérienne. B – Rhizome. C – Racines. D – Radicelles La tige aérienne son organisation : Sans coloration : Coupe colorée ( Acridine /Bleu alcian ) A—Parenchyme de la moelle. B—Cylindre central : Du côté externe phloème et côté interne xylème. C—Gaine de sclérenchyme D—Zone épidermique Aspect sur l’ organisation des cellules de la zone médullaire Coloration en jaune vert avec la Pyranine. Jonction tige aérienne et rhizome : Les tiges aériennes prennent naissance sur une zone du rhizome: Cette photo expose la technique employée par le framboisier pour déclencher la croissance d’une tige aérienne La transformation se fait dès le centre ,donc dans le bois du rhizome , où les méristèmes primaires reprennent du travail ;ces méristèmes développent un phloème qui va permettre la croissance progressive en longueur de la tige La reprise de fonction des méristèmes secondaires vont permettre ensuite à la tige de prendre de l’ épaisseur Le rhizome son organisation Aspect de la coupe.longitudinale Coupe sans colorant.: Coupe axiale Coupe après coloration Acridine / Bleu alcian A – B – C - voir ci-dessous D---Rayons parenchymateux il faut noter ici l’exiguïté de ces rayons : A – Xylème secondaire du bois. B –Phloème secondaire ou liber avec sur son versant interne la région du cambium libéro-ligneux. C –Zone épidermique On constate une organisation radiée où les vaisseaux occupent la presque totalité de l’espace Cette prépondérance des éléments de conduction et de soutien au détriment des tissus parenchymateux contribue à conférer à la structure un caractère de compacité et de robustesse. L’architecture est celle d’une tige secondaire (donc représentative de ce qui se passe dans le tronc des arbres et de ses branches) Les rhizomes sont aussi une zone de stockage: Les Polysaccharides sont mis en évidence sur une coupe traitée avec du Lugol ici localisation des amyloplastes qui apparaissent sous forme de tâches bleu-noires A réserve dans les tissus de soutien B réserve dans les rayons parenchymateux. Comme pour l’ortie ces amyloplastes contiennent de l’Amylose , de l’Amylopectine , de l’ Amidon Les racines De ce rhizomes de développent un réseau racinaire. L’ organisation histologique est la même que celle du rhizome avec un aspect radié et une épaisse couche corticale - Le tissu épithéliale a lui tendance à se détacher facilement ( il correspond au rhytidome : l’ écorce des arbres ) Les radicelles : Ici le parenchyme médullaire est présent ; il s’y trouve déjà de nombreux vaisseaux Ce parenchyme médullaire est entouré par un parenchyme cortical très épais ************************************************************************************************* Eléments complémentaires de phytothérapie (référence 1) Sa culture en Europe remonterait à la fin du Moyen Âge Formes galéniques disponibles Teinture-mère de feuille Macérat glycériné de jeune pousse Jus de fruits Propriétés de la plante Feuille ToniqueAstringen Diurétique Utérotonique, l’extrait aqueux de feuilles de framboisier produit une augmentation de contraction des muscles utérins de souris [4], effet antagonisé par le salbutamol et la nifédipineL’effet pourrait s’inverser en fonction de la concentration et devenir utérorelaxant Antioxydant (fruits et feuilles) [5], [6] La feuille de framboisier rouge a été traditionnellement utilisée pour favoriser le travail des parturientes et pour diminuer la douleur pendant l'accouchement Fruit La raspberry ketone (4-(4-hydroxyphenyl) butan-2-one) du fruit est un composé phénolique aromatique similaire aux structures de la capsaïcine et de la synéphrine, connues pour exercer un effet anti-obésité et moduler le métabolisme lipidique [8], [9] Un extrait de framboise rouge enrichi en polyphénols pourrait protéger le cartilage et moduler l’apparition et la gravité de l’arthrose [10], [11 …Bourgeon Antibactérien utilisé traditionnellement contre la diphtérie ( Corynebacterium diphtheriae), anti-oxydant, cytotoxique sur cellules cancéreuses [13 Anti-inflammatoire et antioxydant [14] Hypogonadisme féminin (aménorrhée, pilosité) de la puberté Ménorragie de la ménopause par hyperplasie endométriale Aménorrhée secondaire des ménopauses précoces iatrogènes Dysménorrhée Présentation des bourgeons Macérat glycériné : 50 gouttes matin et soir Eviter la feuille de framboisier pendant le premier trimestre de la grossesse [16] Référence 1 Wikiphyto Framboisier Dominique .
-
Diatomées indicateurs des pollutions métalliques
Dominique. a répondu à un(e) sujet de Dominique Voisin dans Diatomées (Discussions)
Bonsoir Tes photos sont très bonnes. ( petit problème classique avec la balance des blancs ). La détermination précise de la famille des Naviculaceae est franchement difficile. Elle exige la mesure en longueur et largeur et l'espace entre les stries Amicalement Dominique. -
Diatomées indicateurs des pollutions métalliques
Dominique. a répondu à un(e) sujet de Dominique Voisin dans Diatomées (Discussions)
Bonsoir Nenoeil Les photos fournies par Dominique sont du grand art - mais en explorant tes prélèvements tu dois pouvoir nous présenter quelques diatomée de ta riviére Ce qui serait l' idéale est que tu trouvent des diatomées déformées. Amicalement Dominique ( mais un autre Dominique ) -
Bonsoir Jean -Marie Oui le Tomlov donne d' excellentes images jusqu au grossissement 100. Les trois dernières on été pris au microscope classique. Amicalement Dominique.
-
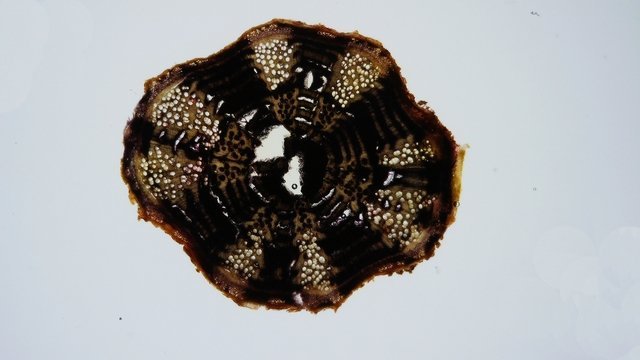
Ortie Rhizome L’ortie a déjà été présentée sur MikrOscOpia. Son système de défense : Son intérêt dans le tissage en raison de la qualité de ses fibres : Reste un point à comprendre - Pourquoi les orties forment parfois des colonies de plusieurs dizaines de mètres carrés si on les laisse faire Quand les orties s’installent dans un coin du jardin il n’est pas facile de les déloger . La raison est simple l’ortie fabrique un rhizome très efficace. Ce rhizome ne court pas très profondément dans le sol - entre 10 et 20 cm de profondeur . En fait les rhizomes les plus puissants sont assez bas dans le sol - les rhizomes de plus petite taille sont eux pas contre à 5 cm sous la surface - Les rhizomes principaux sont souples et très résistants à la fois -Ils se divisent un très grand nombre de fois . C’est sur ces rhizomes que vont se développer au printemps les tiges aeriennes et les racines proprement dit qui elles sont petites et très fines. et de couleur jaune orangée. Aspect macroscopique: Diamètre 1,3 cm. Aspect histologique d’une coupe de rhizome principal. Coloration - Acridine( rouge ) + Bleu Alcian . Ce rhizome est organisé en secteurs avec un fort développement du système vasculaire . Si on regarde ce rhizome en lumière polarisée on constate une importante richesse en fibres ( ce que l’ on trouve sur les tiges florifères et qui en font son intérêt dans l’ industrie textile ). Ces fibres qui expliquent sa résistance et sa souplesse. La disposition de ces fibres est surtout le long des axes vasculaires ; une couche inhomogène se trouve aussi dans le parenchyme cortical. Si on réalise une coloration avec la solution de Lugol on constate la présence de Glucanes qui réagissent avec l’ iode : Dans les racines il peut s’agir d’ d’Amylose , d’Amylopectine , d’ Amidon. Le rhizome est naturellement une zone de réserve .pour la plante . Une étude à fort grossissement met en évidence une forte épaisseur des cellules qui contiennent les réserves .. Axes vasculaires entourés par le parenchyme fait du Xyléme Cette photo permet de constater la très grande épaississeur des parois cellulaires. Par contre les multiples petites racines ont une structure histologique différente: Longueur 7 mm largeur 3 mm . La coupe longitudinale des rhizomes : Cette coupe permet de mettre en évidence l’ organisation de la paroi des vaisseaux. La charge en lignine des fibres du sclérenchyme et des fibres–tracheides n’ est pas homogéne. On le voit ici les épaississements de la paroi secondaire alternent avec des zones composées uniquement de la paroi primaire cellulosique. Cette disposition donne une certaine souplesse au rhizome . Lumière polarisée met en évidence la grande richesse minérale des rhizomes. En lumière normale au 1000: En conclusion pour en savoir plus sur l’ortie - Depuis l’Antiquité l’ortie est considérée comme un hémostatique. Dioscoride la prescrivait pour les métrorragies, les blessures infectées et les saignements de nez. - Avant l’introduction du coton, l’ortie était la plante la plus utilisée en Europe dans la fabrication de vêtements. - Au Moyen Âge, on la trempait dans l’urine d’un malade. Si elle restait verte jour et nuit, elle était le signe d’une guérison imminente, dans le cas contraire, tout espoir était perdu. - Au XVIIIème siècle, on la considérait comme « l’un des remèdes les plus assurés pour le crachement de sang et les hémorragies ». Elle était reconnue pour ses propriétés astringentes, anti diarrhéiques, antidiabétiques et dépuratives Indications modernes . ♦ Voie interne : - Système Immunitaire-Asthénie. - Renforce le système immunitaire. - Traitement des asthénies fonctionnelles, des asthénies liées à un manque de fer et/ou à des carences en minéraux, fatigue chronique, convalescence… - Système articulaire. - Traitement symptomatique des manifestations articulaires douloureuses, arthrite rhumatoïde. - Système génito-urinaire. - Traitement diurétique complémentaire des infections urinaires et des calculs rénaux. - Hyperplasie bégnine de la prostate (racines) : effet décongestionnant et anti-inflammatoire. ♦ Voie externe : Système cutané. - Utilisé dans les états séborrhéiques de la peau. - Traitement d’appoint des affections dermatologiques : dermatoses, eczéma, psoriasis… - Soin des cheveux et du cuir chevelu. Mode d’emploi. ♦ Voie interne : - TM ou extrait fluide : 30 à 40 gouttes 2 à 3 fois / jour. - Gélules, comprimés de poudre de plantes ou d’extraits standardisés. SIPF – EPS. - Tisane : Laisser infuser 30 à 40 g de feuilles d’orties séchées dans 1 litre d’eau pendant 10 min. Filtrer et prendre une tasse avant les repas. ♦ Voie externe : - Lotion capillaire : Faire macérer 60 g de feuilles d’orties avec 60 g d’origan dans un litre d’eau-de-vie pendant un mois. Faire des frictions journalières avec cette lotion. Lotion capillaire contre la chute des cheveux et les pellicules : dans 1 litre d’eau de vie, laisser macérer pendant 15 jours 50 g de racines d’orties concassées, 30 g de feuilles d’orties et 30 g de romarin en prenant soin de bien couvrir le récipient et d’agiter régulièrement. Filtrer. À utiliser chaque matin en frictions sur le cuir chevelu durant environ 2 minutes. ________________________________________ Références: https://fr.modernagriculturefarm.com/plantation/1003014105.html Florence Biarrat Phyto-aromathérapeute Monthey - canton du Valais Suisse Dominique.
-
Le bassin des chironomes est resté dehors - Avec la chute des températures les larves se sont immobilisées dans le fond et sur la paroi. Il y a 1 semaine sont apparus sur la surface de l’eau des chironomes en métamorphose -La forme adulte n’a pas réussi à s’envoler - Ce qui a permis de photographier le moment de leur transition entre la forme larvaire et la forme ailée . ( Il y a eu un petit problème de préparation – le zone thoracique s’ est séparée ). On s’aperçoit que la mue se fait à l’intérieur de la paroi de la larve sans passer par une forme pupe comme le font les mouches ni évidemment par la forme chrysalide comme le font les papillons . En effet, la pupaison se réalise à l'intérieur de la cuticule du dernier stade larvaire, qui n'est pas rejetée, mais qui se durcit et devient un cocon abritant l'ensemble des transformations de la métamorphose. aboutissant à l'imago ,.Dans le cas du chironome cette pupaison semble se faire sans modification des parois de la larve. En biologie, le terme d’imago (au masculin) ou de stade imaginal désigne le stade final d'un individu dont le développement se déroule en plusieurs phases Dominique.
-
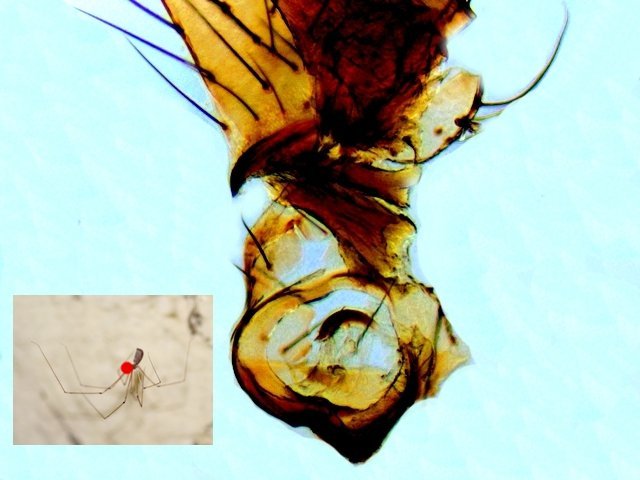
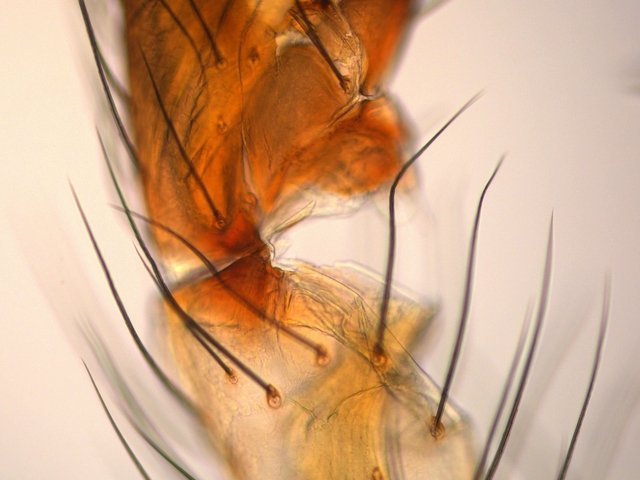
Tomlov Eclairage Le tomlov est vendu avec deux Led assurant un éclairage des sujets par le dessus. Traditionnellement les microscopes sont éclairés grâce à l éclairage dit de Köler Dans le cas de l'illumination de Köhler, l'image de la source est produite dans le plan du diaphragme d'ouverture du condenseur. L'image de la source n'est donc pas superposée à celle de l'échantillon. Ainsi défocalisé, chaque point de la source illumine tout l'objet, rendant l'illumination uniforme. Désormais cette homogénéité de l’ éclairage peut être obtenue avec les Led organisées dans les tablettes lumineuses .Ces tablettes sont normalement faites pour le dessin .. De ce fait leur taille fait qu’elles ne sont pas la solution pour pouvoir se glisser sous l’ objectif . il existe une table lumineuse de petite taille qui remplit parfaitement la demande la Cs-lite camera scanning qui a été développée pour permettre de digitaliser des photos diapositives. . Sa taille 175x120x20 mm . Il existe 3 caches pour limiter le champ d’illumination. https://www.digit-photo.com/CINESTILL-Tablette-Lumineuse-CS-Lite-IRC-95-rCINESTILL69141.htm 39 euros Le Tomlov n’ayant qu’ un éclairage direct épiscopique assuré par deux Led orientables le fond des photos est de ce fait noir. Cette petite table lumineuse corrige efficacement ce manque. L’intensité lumineuse n’est pas réglable ni la température de la lumière mais 3 températures sont qu’en même proposées - En fait le système électronique de ce microscope à écran régule l’éclairage automatiquement Image obtenue Chélicéres de type crochet d’ une petite araignée : Dominique
-
Bonsoir Jean-luc Merci de ton appréciation sur ce sujet . Ce dessin sur le céphalothorax m' apparaît comme une marque qui doit être spécifique de l’espèce mais ce n' est probablement pas un organe vu en transparence. Amicalement Dominique.
-
-
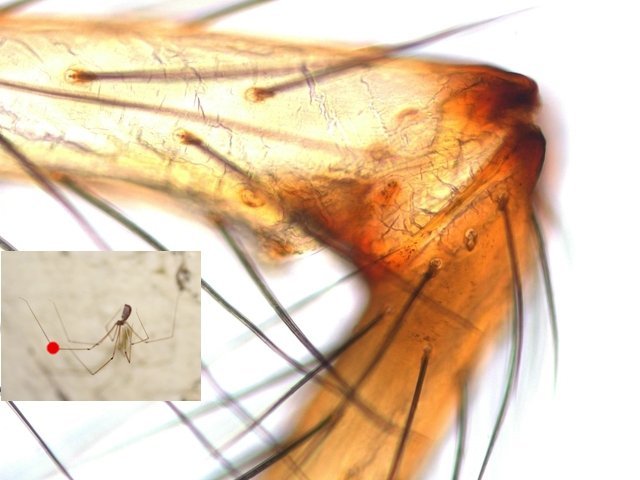
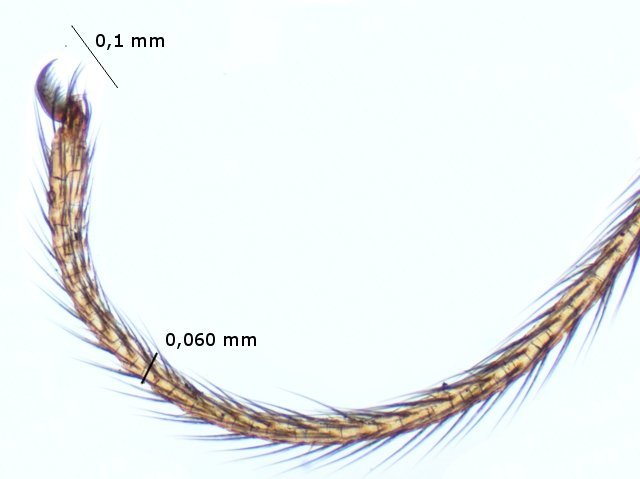
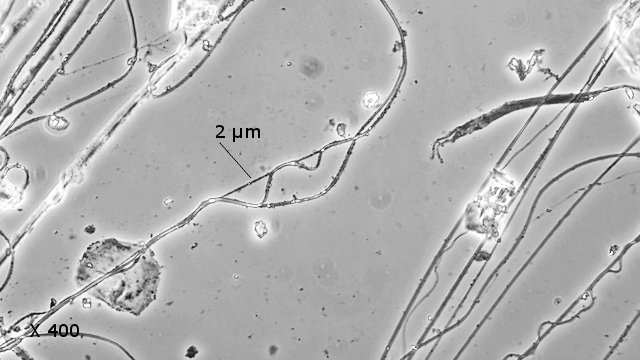
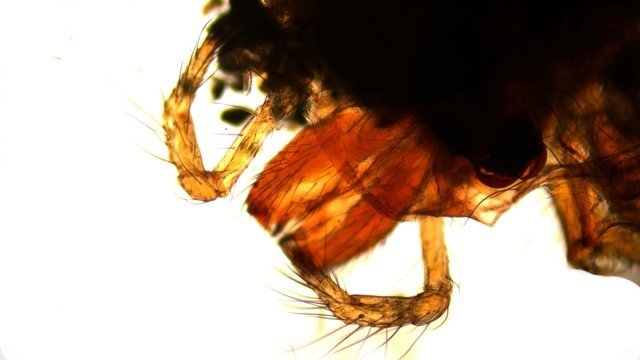
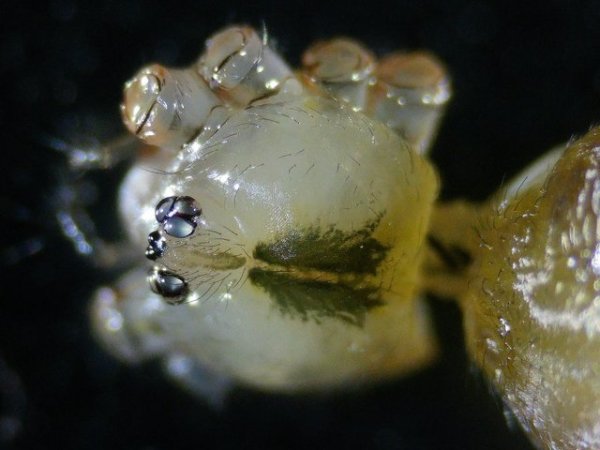
Pholcus phalangioïde Un peu partout dans la maison il existe une petite araignée très discrète que l’ on trouve toujours la tête en bas suspendu à des fils à peine visibles Une partie des images a été obtenue avec le microscope à écran Tomlov . La taille : céphalothorax et corps 6 mm Si l’on touche sa toile, l’araignée se met à tournoyer sur elle-même à grande vitesse au point de ne plus savoir où elle est . : Parfois elle peut se laisser tomber et courir très vite dans un autre petit recoin. Elle peut rester immobile des jours entiers suspendue à sa toile dans l’attente qu un moustique ou une drosophile viennent se coller aux fils de la toile: Les pattes de Pholcus: Ce qui caractéristique cette araignée est la longueur de ses pattes.La Hulotte, nous révèle que mises bout à bout les pattes du Pholcus feraient une longueur de 30 cm soit 40 fois la longueur du corps. . Articulations des pattes: Son nom phallangioïde est bien porté ; L’organisation est la même que les os de nos doigts ( sauf le pouce ) soit 3 segments et 3 articulations Examen depuis la zone d’insertion au céphalothorax vers l’extrémité distale . P1 proximale ( contre le corps ): P2 intermédiaire : la première image étendue , la seconde pliée : Articulation distale P3: La « patte» Les pattes se terminent par une petite griffe qui permet à l ‘araignée de se maintenir sur la toile sans effort, des jours entiers et par un organe constitué de « doigts » qui lui servent à se mouvoir sur les fils de la toile. Examen de l’extrémité distale de la patte qui a développé les éléments d’accroche : 1 griffe qui permet de rester accroché à un fil et un ensemble de doigts qui autorise le déplacement rapide sur les lignes de la toile. La toile Pholcus ne fabrique pas de belles toiles comme on peut en croiser quand on se promène dans la campagne -Ici le but poursuivi est de tendre des haubans en les accrochant sur les éléments porteurs de son environnement . L'araignée possède plusieurs glandes séricigènes, chacune produisant un type de soie avec des propriétés spécifiques (résistance, élasticité, adhésivité...). Lorsqu'elle tisse sa toile, elle utilise ces différentes glandes de manière sélective pour créer les fils adaptés à la fonction qu'ils vont remplir (structure de la toile, capture des proies, enveloppement du cocon...). Le fil de soie de base est d’une grande finesse il est mesuré à 2 µm - le cordage de soutien lui est réalisé par un entortillement de très nombreux fils avec des fibres plus grosses. Le nombre de microfibres est considérable ; elles sont à la base du piège invisible qui assure la capture des proies (-Les fils qui sont visibles à nos yeux sont des câbles souvent multifibres de soutien sur lesquelles se déplace l’ araignée , il y a aussi un grand nombre de microparticules de poussière qui améliorent leur visibilité ) Les petits insectes ne savent pas éviter le piège parfaitement invisible de ces microfibres ,leurs poils vont alors s’ emmêler dans ces fils laissant le temps à l’ araignée de percevoir les vibrations .Elle se précipite pour les attraper et les enrubanner. Pour se faire les pattes arrières se replient jusqu’ aux filières tout au bout de l’abdomen – saisissent un début de fil de soie et tirent ce fil autour de la proie Les deux pattes arrière agissent de concert tandis que les pattes avant permettent le maintien dans la toile de l’araignée. Les poils ; Les poils sont ici tous semblables ;- ils ont un rôle de capteur de vibrations. Les sensilles sont des structures cuticulaires chez les insectes, jouant un rôle sensoriel en détectant des stimuli ;Pour pholcus il est fondamental de percevoir la moindre vibration de la toile Ces sensilles sont montées sur un organe mécanorécepteur le Trichobothrie. Le céphalothorax : La tête et le thorax ne font qu’une unité anatomique: le céphalothorax. Comme éléments caractéristiques de cette araignée il y a les yeux au nombre de 8 2 groupes de 3 et un groupe médian de 2 . Les chelicéres: Les chélicères Les chélicères sont des appendices pairs, les pièces buccales servant à mordre leur proie. Chaque chélicère est un appendice pointu, terminé en croc, utilisé pour saisir la nourriture, et se retrouve à la place d'une mandibule chez la plupart des autres arthropodes . Les chélicères sont situés immédiatement avant la bouche, de sorte qu'ils ne sont pas homologues aux pattes-mâchoires des crustacés, mille-pattes et insectes. Chez les arachnides,les chélicères sont creux et connectés à des glandes à venin; . La digestion se fait en 7 à 8 heures (jusqu’ à 15 h) elle se déroule en deux temps . 1 injection d’enzymes digestives qui dissolvent les organes internes de la proie. 2 aspiration du liquide de digestion. Le reste de l’insecte est rejeté par section des soies qui retenaient le corps désormais vidé de ses organes internes. La reproduction A l’extrémité de l’ abdomen la femelle se trouve l’organe copulatoire : épigyne qui est la pièce tégumentaire génitale externe présente chez certaines femelles d'araignées. Généralement, le mâle maintient la femelle avec ses pattes avant et tente de la soulever suffisamment afin de placer ses bulbes copulateurs en contact avec l'épigyne. La forme de cette plaque correspond à celle des bulbes portés par les pédipalpes du mâle. Extérieurement il n’ y a pas de différence entre le mâle et la femelle en dehors de la transformation des chélicères en deux petits bulbes copulateurs de forme arrondie ( donc le mâle ne peut ,désormais, plus se nourrir ) . Le mâle fabrique le sperme dans son abdomen Ce sperme est pompé par les deux palpes qui deviennent deux organes copulateurs ;ils s’ y remplissent et se transforment en deux petites outres . Le rôle du mâle va alors d’approcher la femelle sans se faire attraper et manger par celle – ci. Il va alors s’accrocher à l’épigyne de la femelle par ses chélicéres et y rester accroché de1 à 3 heures Une fois le transfert de sperme terminé le mâle se laisse tomber au sol à moins que la femelle n’ en ai fait son repas. Il va s’en suivre la ponte de 40 à 60 œufs que la femelle a fixés en une grappe. Cette ci sera maintenue par la femelle entre ses chélicères tout le temps de la gestation ( donc sans s’alimenter ) Elle va ainsi rester 3 semaines le plus souvent immobile suspendue dans sa toile ( parfois elle dépose le fardeau et il peut arriver qu’elle laisse son sac dans la toile pour aller chercher une proie bien venue. A la naissance les jeunes Pholcus sont libérés des fils qui les maintenaient .Ils vont rester immobiles 1 semaine puis subir leur premières mue Ils vont alors se disperser rapidement car - la mère est toujours capable d’en manger quelqu’uns. La quantité de sperme emmagasiné permet à Madame Pholcus de fabriquer 2 à 3 cocons de suite. Mais l’histoire du Pholcus a d’autres aspects :( lire la Hulotte.). Rérérence https://www.lahulotte.fr/ La hulotte Si l’ histoire du Pholcus vous intéresse je vous engage à lire les n° 54 et 55 de ce merveilleux petit journal de sciences naturelles . Dominique.
-
Nouvelles du pouce.
Dominique. a répondu à un(e) sujet de Tryphon T dans Sujets de Microscopes à écran.
Bonsoir Tryphon C'est une belle observation ! Si tu utilises des tissus collants modernes comme Omnifix tu ne risques pas d' asphyxier ton bout de doigt ( avec les sparadraps classiques c'est en effet plus discutable ). La mise en longitudinal est un conseil de grande valeur . Ta dernière image est tout à fait rassurante ; tu dois avoir un excellent terrain vis à vis de la cicatrisation . Amicalement Dominique. -
Nouvelles du pouce.
Dominique. a répondu à un(e) sujet de Tryphon T dans Sujets de Microscopes à écran.
Bonsoir Tryphon Ton ongle est propre. Si tu veux une cicatrisation qui ne traîne pas trop je te conseil de mettre de la Vaseline et de te faire une petite poupée 24h sur 24. La raison est simple l' organisme ne supporte pas la déshydratation - de ce fait il fait une croute quand la peau et ici l'ongle ne réalise plus son rôle de protecteur - Le problème est que sous les croutes la cicatrisation est très ralentie. La vaseline forme une couche protectrice vis à vis de l' oxygène et de la déshydratation .L' épithélium se reforme rapidement ce qui permet d' attendre la lente repousse de l' ongle ( quelques semaines ) le pansement quotidien prévient de l' infection qui est toujours aux aguets. Nettoyage au savon simplement Amicalement Dominique -

-
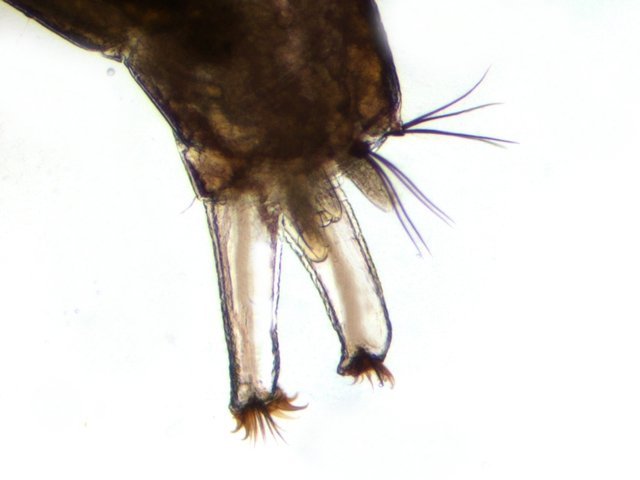
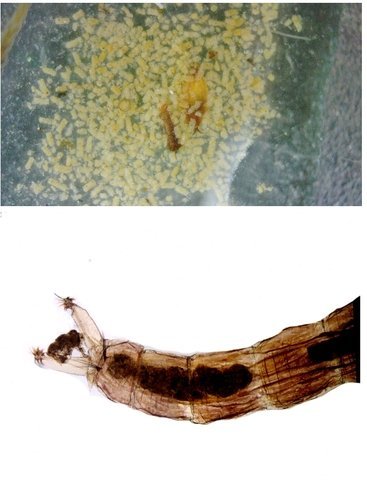
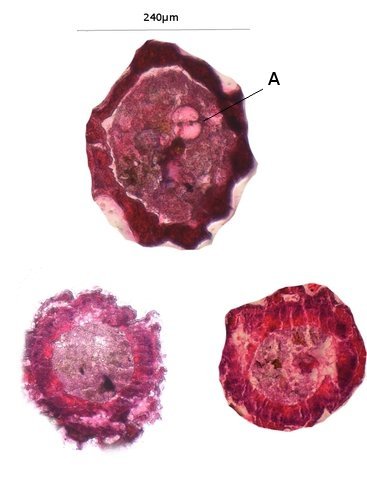
Chironomes A la fin du mois d’Aout devant la maison il y avait tout un nuage de tout petits moucherons .Ce phénomène est bien connu de tous L’ expérience a été de simplement placer sous la zone du vire voltage des moucherons une large boîte transparente remplie d’ eau et de suivre le contenu de cette boite 3 mois. Succession des événements : Constatations : le dépôt de quelques petites feuilles apportées par le vent -Verdissement très progressif de l’eau -Apparition de larves de moustiques -Développement progressif au fond de la boite d’une tache de couleur marron aux contours imprécis -Apparition sur les parois transparentes de la boite de tout petits vers -Prélèvement de quelques-unes de ces petites larves . La boite est alors fermée On découvre une colonie de petits "vers " : Cette larve fait 1, 3 cm. La tête : Vue par en dessous aspect de la mâchoire Les pattes avants : Le corps est fait d’une succession de segments au nombre de 12: La partie postérieure est complexe 1 les pattes arrières Ces pattes arrières ont deux fonctions L’accroche aux éléments environnants Le déplacement par un mouvement de haut en bas comme la nageoire caudale des dauphins. L’ extrémité des pattes arriéres est munie de plusieurs griffes dont le rôle est le maintien sur les supports végétaux L ’anus et la partie terminale de l’intestin: Dans la boite on a vu que le fond était de couleur marron - En fait cette tache est faite de l’accumulation des excréments des vers de chironome. Pour compléter ce petit dossier J’ ai essayé de faire des coupes de cette larve .Les résultats ne sont pas bons -( les tissus périphériques se sont en parti dissociés mais 3 images sont extraites des coupes – épaisseur 7µm – coloration hématoxyline / éosine . L’ image du haut en A montre une coupe de l’ intestin qui semble présenter une membrane sur chaque côté Chez le ver de terre il avait été mis en évidence une membrane dans l’ intestin ;cette membrane a pour but d’ augmente la surface d’ absorption le Typhlosole ref https://forum.MikrOscOpia.com/topic/17385-ver-de-terre-lombric-histologie/#comment-69620 Par ailleurs on est frappé par l’ épaisseur de la paroi musculaire qui forme la périphérie du tube et qui permet ce mouvement d’ondulation des chironomes qui ne semblent pas connaître le repos . *********************** La boite fermée Dans la boîte il y a des larves de moustiques et de chironomes. Les premiers insectes à apparaître ont été les moustiques Dans la semaine qui a suivi l’apparition des moustiques sont apparus de petits moucherons. Ce petit diptère fait 1cm Il se pose sur l’eau grâce à la longueur de ses pattes: Une autre caractéristique est le développement des antennes: Les antennes sont couverte de longs cils (elles sont appellées antennes plumeuses ) Quelle est la nature de ces deux appendices entre les antennes ? -Les diptères ont une seule paire d’antennes or ici il semble que l’ on ait affaire à une seconde paire - l’ aspect extérieur ne ressemble pas à des palpes - Au passage on note que la bouche n’est pourvue ni de trompe ni d’ organe piqueur L image ci-dessous donne plus de détails : Image supplémentaire de la partie arrière du chironome le système ovipositeur. ************************* Texte extrait de Wikipedia Les chironomidés sont de petits insectes volants qui appartiennent à la famille des diptères. Ils sont souvent confondus avec les moustiques en raison de leur apparence similaire, mais ils en diffèrent par quelques caractéristiques importantes. Voici quelques points clés pour mieux comprendre les chironomidés : Apparence: Taille: Généralement plus petits que les moustiques, mais leur taille peut varier. Couleur: Souvent de couleur verdâtre, mais cela peut varier selon les espèces. Corps: Fin et délicat, avec de longues pattes frêles et des ailes fragiles. Absence de trompe: Contrairement aux moustiques, ils n'ont pas de trompe pour piquer. Comportement: Non-piqueur: Les chironomidés ne piquent pas les humains ou les animaux. Leur nom anglais "non-biting midges" signifie d'ailleurs "moucheron non-piqueur". Milieu de vie: On les trouve souvent près des plans d'eau douce, comme les lacs, les rivières et les étangs, car leurs larves se développent dans l'eau. Activité: Ils sont particulièrement actifs lors des journées chaudes et ensoleillées. Rôle écologique: Chaîne alimentaire: Les larves de chironomidés constituent une source de nourriture importante pour de nombreux poissons et autres animaux aquatiques. Bioindicateurs: La présence de certaines espèces de chironomidés peut être utilisée pour évaluer la qualité de l'eau. En résumé, les chironomidés sont des insectes inoffensifs qui jouent un rôle important dans les écosystèmes aquatiques. Bien qu'ils puissent être présents en grand nombre et former des essaims, ils ne représentent aucune menace pour la santé humaine. Reference https://fr.wikipedia.org/wiki/Larve_de_chironome https://forum.MikrOscOpia.com/topic/15106-le-moustique-tigre/#comment-56544 reflexion de solito de solis sur le sujet des chirnomes https://forum.MikrOscOpia.com/topic/13657-chromosomes/#comment-47988 dissection par Pierre et coloration des glandes salivaires d’un ver de vase qui sont des chironomes rouges le plus souvent et de taille supérieure à celui de la présentation. Dominique.
-
insecte 5.jpg
Dominique. a posté une image de la galerie dans Microscopes à écran. de Microscopes à écran.

-
Bonsoir Tryphon Merci pour le protocole. On va mettre cela en fonction avec le prochaine article Amicalement Dominique
-
Bonsoir Jean-Marie Le problème avec le microscope à écran est que le grossissement ne peut être calculé ( c"est toujours possible mais il y a beaucoup de manipulation ). disons au doigt mouillé que l'on se trouve entre x40 et x 80 . Les glandes sécrétrices sont visible à l' oeil nu . Amicalement Dominique.
-
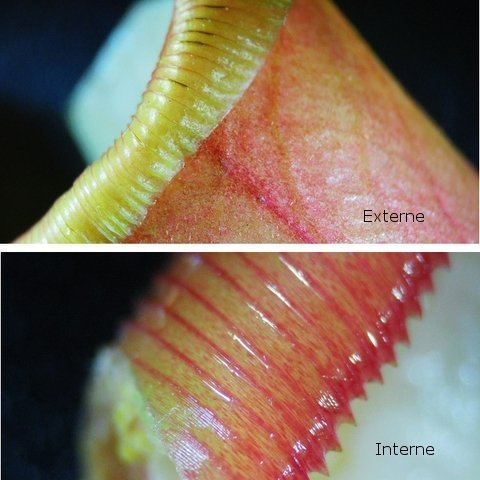
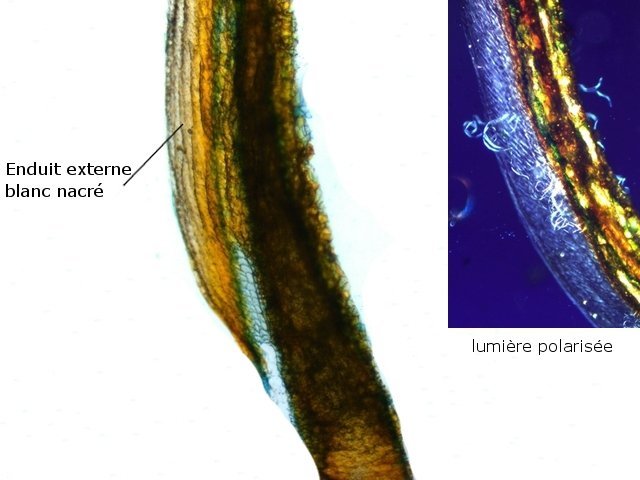
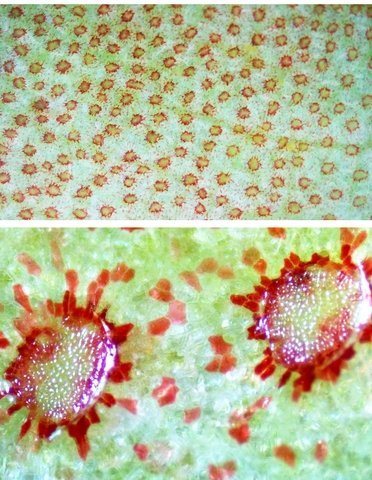
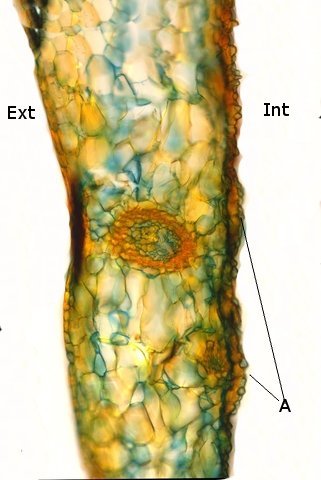
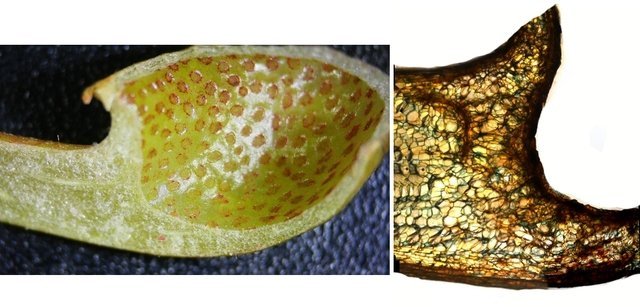
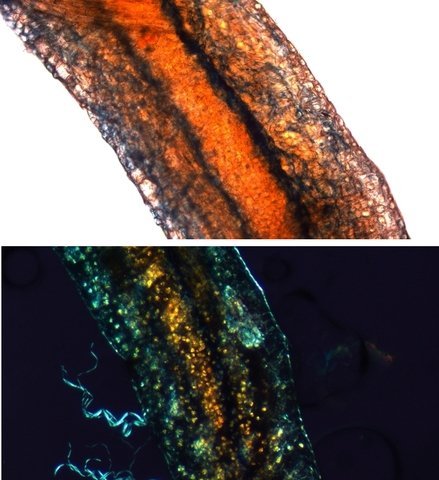
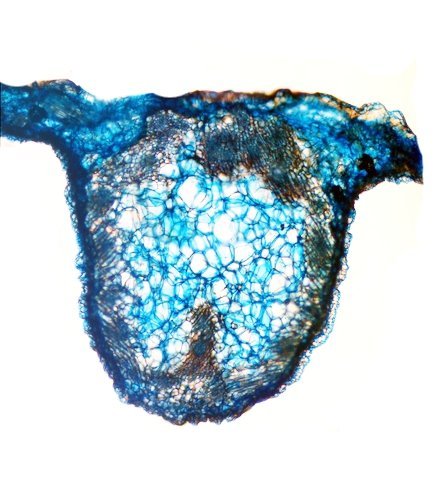
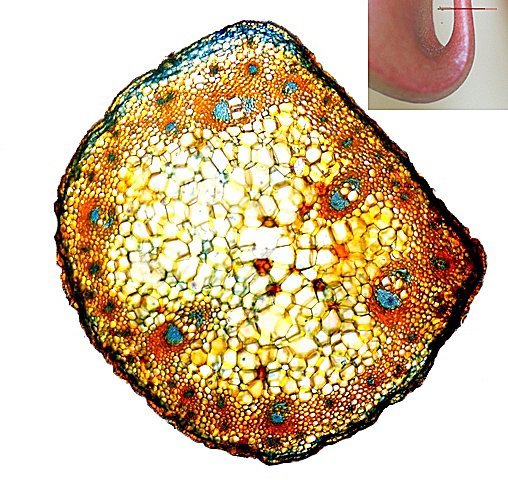
Népenthes alata plante carnivore Il m’a été offert une plante à l’aspect inhabituel : Cette plante qualifiée de carnivore est intéressante à analyser. On constate que les pièges sous forme d’urnes sont en fait le développement de l’extrémité distale des feuilles ( on verra plus tard l’ intérêt de cette disposition )..et de ce fait ce sont les premiers éléments de la feuille à apparaître lors de la croissance de la plante . Feuille à un stade précoce de son développement . ( taille à ce moment 2cm ). Une fois le piège formé le développement de la feuille se fait sur le mode classique avec une nervure centrale et deux limbes Le piège Il est formé par une urne membraneuse ou ascidie ( définition : 1. Organe ayant la forme d'un récipient, parfois muni d'un opercule, et situé à l'extrémité des feuilles de certains végétaux :surmontée d'un opercule – A ne pas confondre avec l’acidie en relation avec les tuniciers qui sont des animaux marins ) .L’ analyse de l’ouverture de l’ascidie met en évidence un système chimique d’attraction des insectes Si on regarde cette ouverture elle est surmontée d’une sorte de couvercle ouvert La paroi de ce couvercle présente un enduit blanchâtre visible à la binoculaire mais pas vraiment visible à l’ œil nu. C’est un enduit poussiéreux dont aucune odeur ne se dégage du moins pour un nez humain . La bouche de l’ urne a deux pans différents : Sur une coupe microscopique on distingue : L’enduit est positionné sur le couvercle mais aussi sur le versant externe du bord de l’ urne . Les ascidies, qui sont les pièges de la plante, renferment un peu de liquide très faiblement sucré et à peine acidulé sécrété par leurs parois Le PH du milieu est de 6,5 Les glandes sécrétrices sont bien visibles sur la face interne de l’ urne. Une coupe de la paroi de cette urne est réalisée. En A se trouve les clusters des cellules synthétisant les sucs digestifs L’ urne est située au bout de la tige qui la relie aux limbes de la feuille ( en fait une extension de la nervure principale de la feuille ) Image de la division de la tige pour former l’ urne: La tige qui forme l’urne est due au remaniement génétiquement déterminé de la nervure principale de la feuille à son extrémité distale. Coupe de cette tige : Ces tiges ont une quinzaine de centimètres de long .Au sommet de celles-ci les deux limbes de la feuille se développent avec une grosse nervure centrale. Coupe d’un limbe de feuille : L’organisation de cette feuille est particulière il n’ existe pas la distinction entre le parenchyme pyramidal lieu de la photosynthèse et le parenchyme lacuneux lieu des échanges gazeux Dans le cas de la feuille de la Nepenthes l’ organisation semble symétrique avec une nervure centrale. En lumière. polarisée on constate une forte charge de corps biréfringents .La mastication de cette feuille révélè une légère amertume est un picotement sur les lèvres . qui va persister une bonne demi-heure (agent chimique ? ) Image de la nervure principale de la feuille : La fonction du piège : Nepenthes est une plante dite carnivore Si on ouvre l’urne --la paroi est humide et le fond de l’urne contient quelques millimètres d’eau seulement. Au toucher la paroi est humide non visqueuse (donc les insectes ne s’y noient probablement pas). -- il est mis en évidence un bon nombre de restes de mouches et de petits papillons et cela dans toutes les urnes. Les corps des insectes ne sont plus reconnaissables Mais il y a une surprise: Cette surprise nous attend ; bien que ‘l’ambiance local soit celle d’une nécropole avec des restes d’insectes bien digérés il existe cependant de la vie . Sur les reste des mouches se promènent en toute impunité des acariens qui ne semblent pas être affectés par les sucs digestifs produits par les glandes digestives de l’ urne Evolution L’urne va tomber aux bout d’un certain temps en se desséchant – le desséchement commence toujours à hauteur de l’ ouverture de l’ urne et la feuille, elle, garde encore un la tige de support de l’ urne .Du moins c’est ce qui se passe à la maison Cette plante est très loin de son lieu d’ origine Elle se développe dans des forêts de nuages, à une altitude variant entre 800 et 2400 mètres au-dessus du niveau de la me Le climat est tropical humide, avec des nuits d’autant plus fraîches que l’altitude est élevée. Le brouillard est omniprésent. Donc dans la cuisine on ne rassemble aucune de ces conditions Donc l' évolution des urnes en milieu naturel doit être différent. . Le piège a donc fonctionné le temps de la croissance de la feuille en apportant à cette croissance des aliments nutritifs complémentaires que les racines ne pouvaient pas apporter Reference : https://www.plantecarnivore.fr/especes/nepenthes/nepenthes-alata/ Dominique
-
Bonjour Tryphon Merci pour le nom et ton complément d' information - La fabrication du vinaigre se fait depuis des générations dans la famille ; on perpétue le rite . J' ai ajouté le nom de ce ver dans le titre et dans l 'article . Amicalement Dominique
-
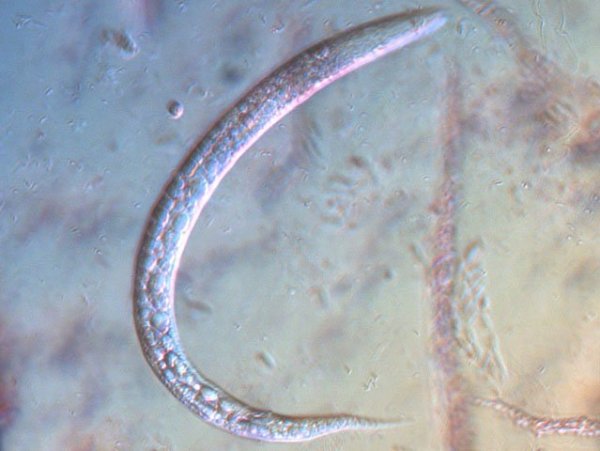
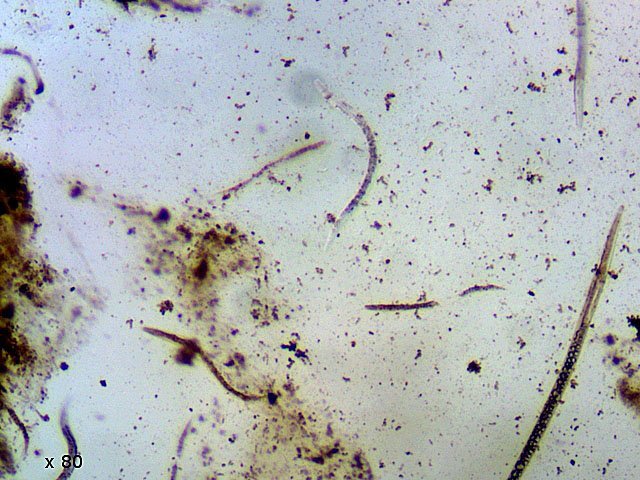
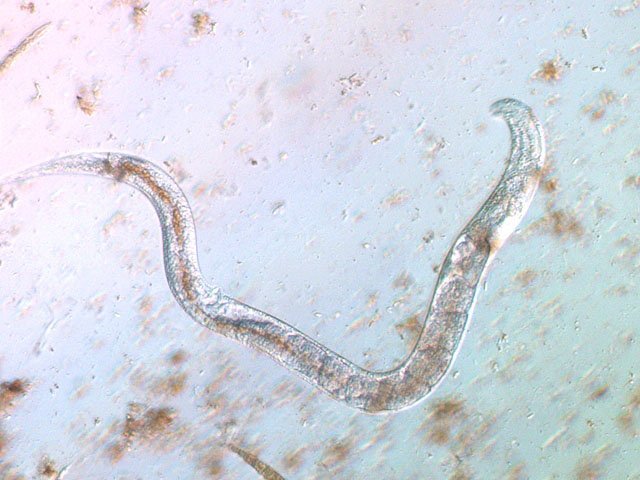
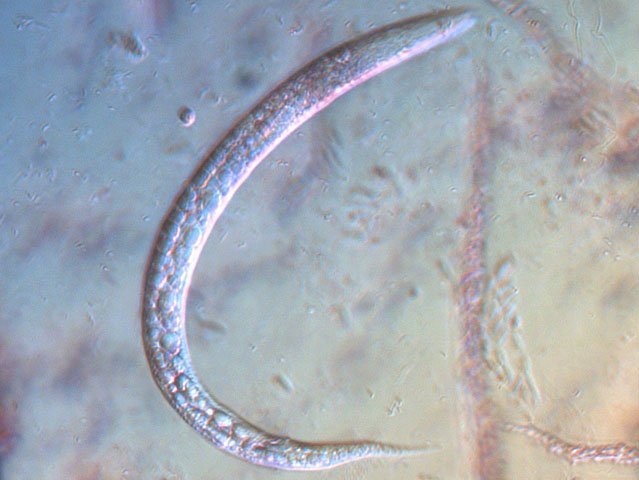
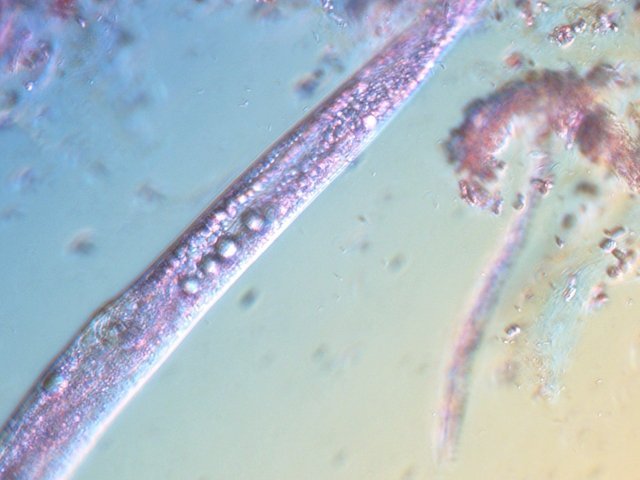
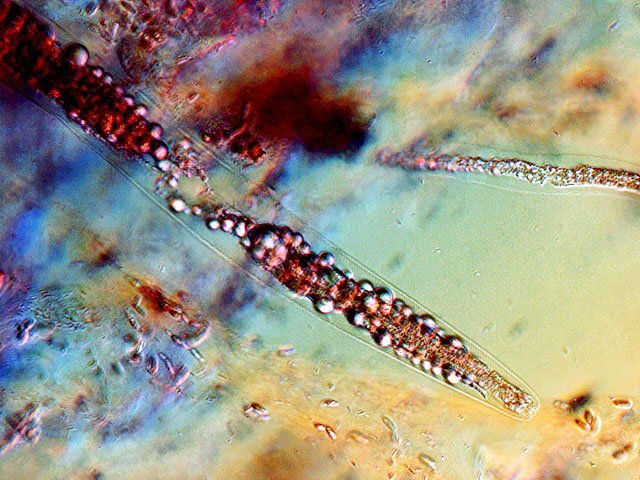
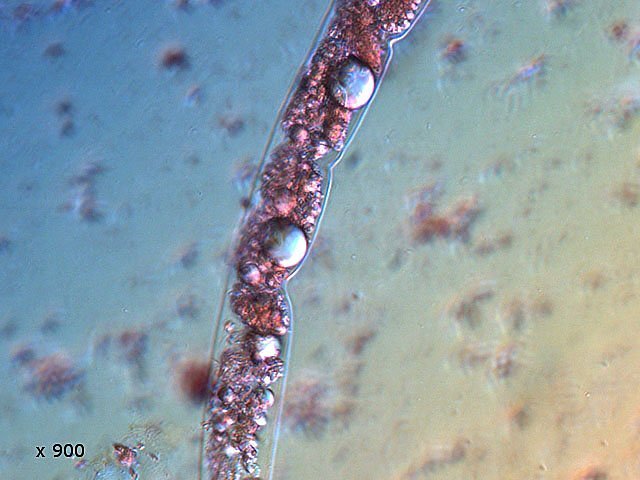
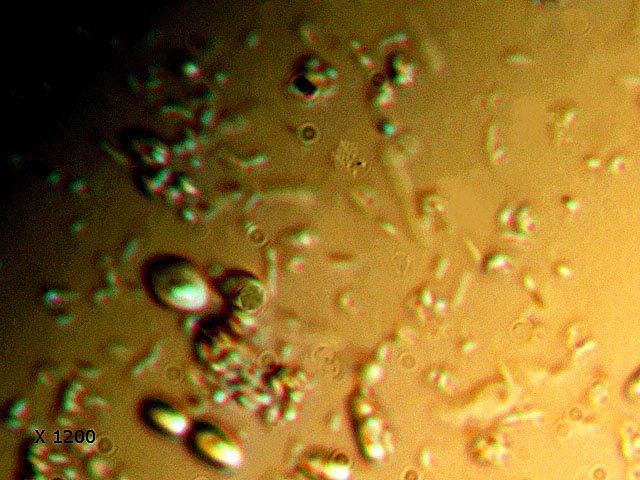
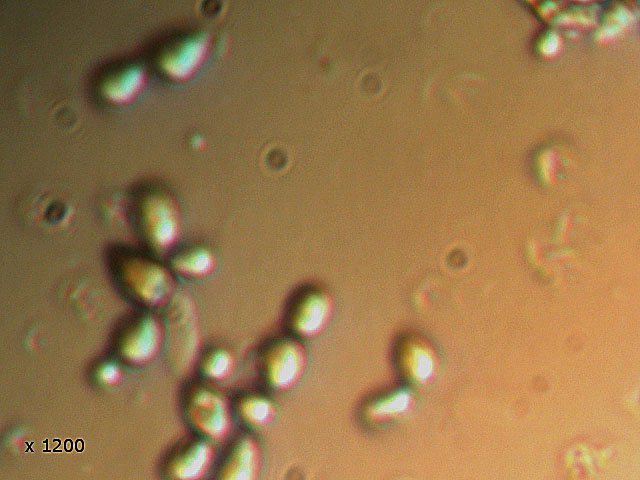
-
Vinaigre A la maison , et cela depuis des années, mon épouse fabrique le vinaigre avec les restes de bouteilles de vin Le vinaigrier est dans la cave . Quand on ouvre le couvercle on constate regardant à l’ intérieur du vinaigrier : Sur la surface et en périphérie existe un dépôt blanc- rosé : dépôt appelé la mère du vinaigre La question qu’un microscopiste se pose est de quoi est formée cette mère . 4 Dans la boite de Pétri la mère est bien distincte du vinaigre sous-jacent. La structure est hétérogène - de type feutrage ; la couleur est liée au vin rouge qui sert à faire du vinaigre. Le Ph du milieu à 20°C est de 2,3 ( soit le ph du jus de citron ) donc milieu fortement acide. Le prélèvement de ce feutrage étalé sur une lame révèle ce à quoi on ne s’ attendait pas ( toute les photos sont prises en Dic ). Dans ce feutrage s’est donc développée une colonie d’un petits vers de 350 µm le milieu est pourtant franchement acide. Il ' agit un bon milieu de conservation si on tient compte du grand nombre de vers morts . Le point curieux est que certains vers semblent contenir des bulles. Ce phénomène s’ exagère énormément dès que le ver est mort. Les vers sous cette forme sont très nombreux dans le milieu. Son nom ( fournit par Tryphon ) est Turbatrix aceti ou Anguillula aceti - La question est comment sont-ils arrivés dans le vinaigrier ? On peut formuler deux hypothèses : --Les œufs ont été apportés par les mouches du vinaigre ((Drosophila melanogaste) qui voltigent autour du vinaigrier et qui y pénètrent par les zones d’aération. --Les œufs ont été importés lors des manipulations pour fabriquer une mère (au départ du processus il avait été mis de petits morceaux de pain dans le vin). IL existe aussi dans cette mère de grandes quantités de levures et de bactéries. Les espèces de bactéries acétiques intervenant dans le processus d'acescence (le nom du processus de formation du vinaigre à partir d’une boisson alcoolisée ) sont principalement : Acetobacter aceti, Gluconobacter oxydans, Acetobacter orleanensis, Acetobacter œni et Gluconoacetobacter europaeus dont la tolérance à l'éthanol et à l'acide acétique est la plus forte. Ces bactéries vont former le film biologique qui est appelé mère . Il existe aussi d’ autres structures totalement immobiles et de grande taille . Je n’ai trouvé aucune information sur ce type de structure dans le vinaigre. Enfin il faut noter l’absence d’éléments mycéliens. Donc la mère du vinaigre est un film biologique formé par la prolifération de bactéries et de levure mais elle peut être aussi être le lieu de développement de certains microvers sans oublier la drosophile dont les pupes restent collées sur le couvercle du vinaigrier. Référence https://fr.wikipedia.org/wiki/Vinaigre#Contamination_fr%C3%A9quente_par_le_plomb_de_certains_vinaigres https://fr.wikipedia.org/wiki/Anguillule Dominique ( NB la présence des petits vers n’’ empêche pas la consommation de ce vinaigre maison dont le goût est très agréable )